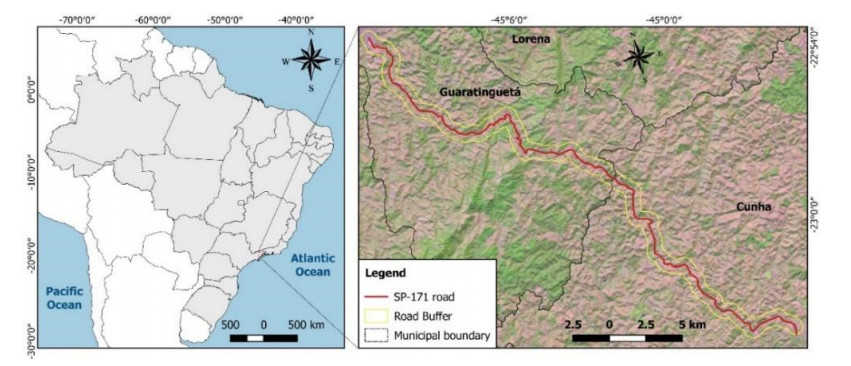
Mass movement susceptibility mapping from rainfall data and in situ site characterization constitute an important approach for preventing geological-geotechnical accidents on railroads and highways. A comprehensive site characterization program was conducted to identify slopes with mass movements along the 44 km of SP-171 road in the state of Sã o Paulo, Brazil. Ninety-two slopes with some degree of instability were found along this section of the road, including rupture scars, active erosive processes and the presence of unstable rock blocks. Two scenarios for mass movement susceptibility (100 mm and 500 mm of accumulated rainfall) were defined by overlaying thematic maps of relief, soil type, geology, accumulated rainfall and declivity using geographic information system-based techniques. The results for both scenarios identified the regions with high and medium susceptibility to mass movements; for the scenario of 100 mm of accumulated rainfall; we found that 27% and 73% of the land area of SP-171 is respectively highly and moderately susceptible to landslide events. For the scenario of 500 mm, we found 58% and 40% to be highly and moderately susceptible areas. This study also allowed us to identify the main geotechnical problems along the 44 km of this road, and thus can be used to guide actions and decisions to avoid or minimize such problems.
Citation: Leila Maria Ramos, Thiago Bazzan, Mariana Ferreira Benessiuti Motta, George de Paula Bernardes, Heraldo Luiz Giacheti. Landslide susceptibility mapping based on rainfall scenarios: a case study from Sao Paulo in Brazil[J]. AIMS Geosciences, 2022, 8(3): 438-451. doi: 10.3934/geosci.2022024
Mass movement susceptibility mapping from rainfall data and in situ site characterization constitute an important approach for preventing geological-geotechnical accidents on railroads and highways. A comprehensive site characterization program was conducted to identify slopes with mass movements along the 44 km of SP-171 road in the state of Sã o Paulo, Brazil. Ninety-two slopes with some degree of instability were found along this section of the road, including rupture scars, active erosive processes and the presence of unstable rock blocks. Two scenarios for mass movement susceptibility (100 mm and 500 mm of accumulated rainfall) were defined by overlaying thematic maps of relief, soil type, geology, accumulated rainfall and declivity using geographic information system-based techniques. The results for both scenarios identified the regions with high and medium susceptibility to mass movements; for the scenario of 100 mm of accumulated rainfall; we found that 27% and 73% of the land area of SP-171 is respectively highly and moderately susceptible to landslide events. For the scenario of 500 mm, we found 58% and 40% to be highly and moderately susceptible areas. This study also allowed us to identify the main geotechnical problems along the 44 km of this road, and thus can be used to guide actions and decisions to avoid or minimize such problems.

| [1] |
Ngo PTT, Panahi M, Khosravi K, et al. (2021) Evaluation of deep learning algorithms for national scale landslide susceptibility mapping of Iran. Geosci Front 12: 505–519. https://doi.org/10.1016/j.gsf.2020.06.013 doi: 10.1016/j.gsf.2020.06.013

|
| [2] |
Azarafza M, Ghazifard A, Akgün H, et al. (2018) Landslide susceptibility assessment of South Pars Special Zone, southwest Iran. Environ Earth Sci 77: 1–29. https://doi.org/10.1007/s12665-018-7978-1 doi: 10.1007/s12665-018-7978-1
|
| [3] |
Ayalew L, Yamagishi H (2005) The application of GIS-based logistic regression for landslide susceptibility mapping in the Kakuda-Yahiko Mountains, Central Japan. Geomorphology 65: 15–31. https://doi.org/10.1016/j.geomorph.2004.06.010 doi: 10.1016/j.geomorph.2004.06.010
|
| [4] |
Nanehkaran YA, Mao Y, Azarafza M, et al. (2021) Fuzzy-based multiple decision method for landslide susceptibility and hazard assessment: A case study of Tabriz, Iran. Geomech Eng 24: 407–418. https://doi.org/10.12989/gae.2021.24.5.407 doi: 10.12989/gae.2021.24.5.407
|
| [5] |
Wu Y, Ke Y, Chen Z, et al. (2020) Application of alternating decision tree with AdaBoost and bagging ensembles for landslide susceptibility mapping. Catena 187: 104396. https://doi.org/10.1016/j.catena.2019.104396 doi: 10.1016/j.catena.2019.104396
|
| [6] |
Guo Z, Shi Y, Huang F, et al. (2021) Landslide susceptibility zonation method based on C5.0 decision tree and K-means cluster algorithms to improve the efficiency of risk management. Geoscience Frontiers 12: 101249. https://doi.org/10.1016/j.gsf.2021.101249 doi: 10.1016/j.gsf.2021.101249
|
| [7] | Departamento de Estradas de Rodagem-DER (2017) Parecer técnico—processo nº 000427/07/der/2.017. Engenheiro Hiromiti Nakao CEE/de – projetos, "In press". |
| [8] |
Tajudin N, Ya'acob N, Mohd ali D, et al. (2020) Estimation of TRMM rainfall for landslide occurrences based on rainfall threshold analysis. Int J Electr Comput Eng 10: 3208–3215. https://doi.org/10.11591/ijece.v10i3.pp3208-3215 doi: 10.11591/ijece.v10i3.pp3208-3215
|
| [9] | Pereira JO (1998) Avaliação das condições dos taludes ao longo da rodovia SP - 171 que liga as cidades de Guaratinguetá e Cunha/SP. Relatório das atividades desenvolvidas. UNESP - Guaratinguetá., "In press". |
| [10] | Arrais LS (2000) Estudo geológico e geotécnico dos solos residuais do pré-cambriano da região da rodovia SP-171. Monografia (Graduação em Engenharia Civil). Universidade Estadual Paulista, Guaratinguetá/SP, 2000, "In press". |
| [11] | Instituto Geológico—IG (2010) Relatório de atendimento técnico—PPDC operação 2009/2010. Decreto nº 42585 de 01.12.97 que dispõe sobre o plano preventivo da defesa civil. Available from: https://www.infraestruturameioambiente.sp.gov.br/institutogeologico?s=cunha. |
| [12] | Companhia de Pesquisa de Recursos Minerais—CPRM (2017) Relatório de ação emergencial para delimitação de áreas em alto e muito alto risco a enchentes, inundações e movimentos de massa, Cunha/SP. Available from: https://rigeo.cprm.gov.br/handle/doc/20711. |
| [13] | Instituto de Pesquisas Tecnológicas—IPT (2018) Reavaliação do mapeamento das áreas de risco no município de Cunha/SP. Relatório técnico nº 154.612-205 SDECTI/PATEM Prefeitura Municipal de Cunha. |
| [14] |
Valladares Soares P, Yoshinaga Pereira S, Coelho Simões SJ, et al. (2008) Mapa de infiltração do alto e médio vale do paraíba do sul com base em elementos do meio físico e na precipitação. Revista Ambiente Água 3: 36–42. https://doi.org/10.4136/ambi-agua.40 doi: 10.4136/ambi-agua.40
|
| [15] | Coelho JOM, Zaine JE, Rodrigues FH (2016) Análise fisiográfica, a partir de técnicas de fotointerpretação, aplicada ao mapeamento geológico-geotécnico de obras rodoviárias. Revista Brasileira de Cartografia 68. Available from: http://www.seer.ufu.br/index.php/revistabrasileiracartografia/article/view/44302. |
| [16] | Bazzan T (2018) Análise da suscetibilidade a escorregamentos no município de Guaratinguetá/SP. Monografia (curso de introdução ao geoprocessamento). Instituto Nacional de Pesquisas Espaciais, São José dos Campos. Available from: http://wiki.dpi.inpe.br/lib/exe/fetch.php?media=ser300:trabalho_final_thiago_bazzan.pdf. |
| [17] |
Ribeiro AJA, da Silva CAU, Barroso SHA (2018) Metodologia de baixo custo para mapeamento geotécnico aplicado à pavimentação. Transportes 26: 84–100. https://doi.org/10.14295/transportes.v26i2.1491 doi: 10.14295/transportes.v26i2.1491
|
| [18] | Instituto de Pesquisas Tecnológicas—IPT (2015) Carta de suscetibilidade a movimentos gravitacionais de massa e inundações: município de Guaratinguetá—SP. 1 mapa, color. Escala 1: 50.000. Available from: http://rigeo.cprm.gov.br/xmlui/handle/doc/15176. |
| [19] | Instituto de Pesquisas Tecnológicas—IPT (2015) Carta de suscetibilidade a movimentos gravitacionais de massa e inundações: município de Cunha/SP. 1 mapa, color. Escala 1: 75.000. Available from: http://rigeo.cprm.gov.br/jspui/handle/doc/15169. |
| [20] | Bitar OY (2014) Cartas de suscetibilidade a movimentos gravitacionais de massa e inundações: 1: 25.000 (livro eletrônico): nota técnica explicativa/coordenação Omar Yazbek Bitar. São Paulo: IPT—Instituto de Pesquisas Tecnológicas do Estado de São Paulo; Brasília, DF: CPRM—Serviço Geológico. |
| [21] | Instituto de Pesquisas Tecnológicas—IPT (2010) Parecer técnico, n. 18.745 - 301 - i/122, 2010. |
| [22] | Simões SJC, Bernardes GP, Trannin ICB, et al. (2012) Relação do meio físico com o manejo sustentável de plantio de eucalipto em áreas de terrenos acidentados—a fazenda Santa Edwirges, Vale do Paraíba, sudeste do Brasil. Geosciences 31: 431–445. |
| [23] | Empresa Brasileira de Pesquisa Agropecuária—EMBRAPA (2013) Banco de Dados Climáticos do Brasil. Available from: https://www.cnpm.embrapa.br/projetos/bdclima/index.html. |
| [24] |
Rozante JR, Moreira DS, Gonçalves LGG, et al. (2010) Combining TRMM and surface observations of precipitation: technique and validation over South America. Weather Forecast 25: 885–894. https://doi.org/10.1175/2010WAF2222325.1 doi: 10.1175/2010WAF2222325.1
|
| [25] | Santoro J, Mendes RM, Pressinotti MMN, et al. (2008) Correlação entre chuvas e deslizamentos ocorridos durante a operação do plano preventivo de defesa civil em São Paulo. Simpósio Brasileiro de Cartografia Geotécnica e Geoambiental. Available from: <https://www.researchgate.net/publication/310801390_correlacao_entre_chuvas_e_deslizamentos_ocorridos_durante_a_operacao_do_plano_preventivo_de_defesa_civil_em_sao_paulo_sp/link/5838371308aef00f3bf9e94c/download. |
| [26] | Alaska Satellite Facility—ASF/Distributed active archive centers—DAAC (2020) ALOS PALSAR Hi-Res terrain corrected. Available from: https://search.asf.alaska.edu. |
| [27] | Departamento de Águas e Energia Elétrica—DAEE (1984) Mapa geológico na escala original 1: 250 000. Universidade Estadual Paulista—UNESP. |
| [28] | Oliveira JB, Camargo MN, Rossi M, et al. (1999) Mapa pedológico do estado de São Paulo: legenda expandida escala 1: 500.000. Campinas: Instituto Agronômico/EMBRAPA solos. |
| [29] | Companhia de Pesquisa de Recursos Minerais—CPRM (2020) Cartas de suscetibilidade a movimentos gravitacionais de massa e inundações—São Paulo. Available from: http://www.cprm.gov.br/publique/gestao-territorial/prevencao-de-desastres/cartas-de-suscetibilidade-a-movimentos-gravitacionais-de-massa-e-inundacoes---sao-paulo-5088.html. |
| [30] | Ramos LM, Bernardes GP, Motta MFB, et al. (2019) Retroanálise da ruptura de um talude rodoviário na SP-171: Cunha/SP. Ⅸ Congresso Brasileiro de Geotecnia Ambiental. Ⅷ Congresso Brasileiro de Geossintéticos. São Carlos, São Paulo, Brasil© IGS-Brasil/ABMS. Available from: http://www.abms.com.br/links/bibliotecavirtual/Regeo-Geossinteticos%202019/Anais_Geossinteticos_REGEO2019.pdf. |
| [31] |
Antunes F, Campos T, Polivanov H, et al. (2013) Desenvolvimento de classes e unidades geo-pedológicas a partir da interação entre a pedologia e a geotecnia. Geotecnia, 61–79. https://doi.org/10.14195/2184-8394_127_3 doi: 10.14195/2184-8394_127_3
|
| [32] | Silva VCB, Machado PS (2014) SIG na análise ambiental: susceptibilidade erosiva da bacia hidrográfica do córrego mutuca, Nova Lima—Minas Gerais. Revista de Geografia (UFPE). Available from: https://periodicos.ufpe.br/revistas/revistageografia/article/view/229090. |
| [33] | Prefeitura Municipal de Angra dos Reis/RJ (2010) Prefeitura preparou o formulário AVADAN. Available from: https://www.angra.rj.gov.br/noticia.asp?vid_noticia=30386&indexsigla=imp. |
| [34] | Gerscovich DMS (2016) Estabilidade de taludes. 2 ed., São Paulo: Oficina de textos. |
| [35] |
Di B, Stamatopoulos CA, Stamatopoulos AC, et al. (2021) Proposal, application and partial validation of a simplified expression evaluating the stability of sandy slopes under rainfall conditions. Geomorphology 395: 107966. https://doi.org/10.1016/j.geomorph.2021.107966 doi: 10.1016/j.geomorph.2021.107966
|
| [36] |
Take WA, Bolton MD, Wong PCP, et al. (2004) Evaluation of landslide triggering mechanisms in model fill slopes. Landslides 1: 173–184. https://doi.org/10.1007/s10346-004-0025-1 doi: 10.1007/s10346-004-0025-1
|
| [37] |
Huang Y, Zhao L (2018) Review on landslide susceptibility mapping using support vector machines. Catena 165: 520–529. https://doi.org/10.1016/j.catena.2018.03.003 doi: 10.1016/j.catena.2018.03.003
|
| [38] |
Chen X, Chen W (2021) GIS-based landslide susceptibility assessment using optimized hybrid machine learning methods. Catena 196: 104833. https://doi.org/10.1016/j.catena.2020.104833 doi: 10.1016/j.catena.2020.104833
|





Figures(6) / Tables(2)

Leila Maria Ramos, Thiago Bazzan, Mariana Ferreira Benessiuti Motta, George de Paula Bernardes, Heraldo Luiz Giacheti. Landslide susceptibility mapping based on rainfall scenarios: a case study from Sao Paulo in Brazil[J]. AIMS Geosciences, 2022, 8(3): 438-451. doi: 10.3934/geosci.2022024







 DownLoad:
DownLoad: