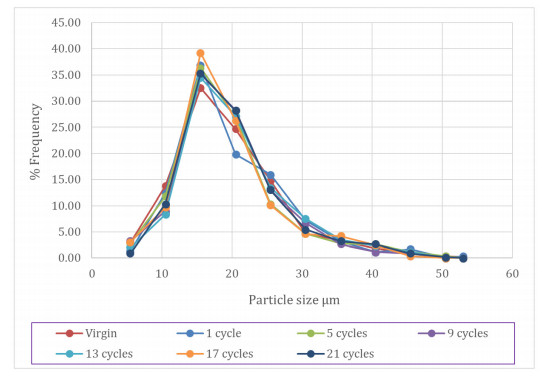
Additive manufacturing technologies such as laser powder bed fusion (LPBF) provide unprecedented abilities to manufacture complex 3-D parts and structures; however, the process produces leftover metal powder that is often unused, resulting in significant cost and disposal overhead. Towards this end, to ensure the ability to recycle the unused powder that is typically found within the build volume and in the overflow compartments of LPBF systems, a thorough chemical and structural analysis is required. Such an analysis will help in determining the extent of recyclability and reusability of the unused powder. For these purposes, characterization of recycled powder for Inconel 718 was performed in this work as a function of build-cycle to compare and contrast the evolution in physical and chemical properties such as particle size distribution, morphology, and composition. We note that the analysis was carried out for 21 build-cycles. Characterization of the particle size distribution and circularity was performed using traditional scanning electron microscopy methods and that information was later compared to measurements of surface profile using coherent scanning interferometry. Reusability was assessed using these results in conjunction with chemical analysis of the powder that was carried out using energy dispersive X-ray spectroscopy. For the extent of build-cycles considered in this work, it was seen that the physical and chemical changes in the recycled powder were will within accepted limits for powder usage and that with appropriate measures, recycled powder can be re-incorporated for subsequent builds without compromise in the powder quality. This study provides a pathway for greatly reducing powder consumption and thus cost during the LPBF process.
Citation: Susana J. Castillo, Anna Hayes, Greg Colvin, Barrett G. Potter, Rongguang Liang, Krishna Muralidharan. Characterization of recycled Inconel 718 metal powder for assessing its reusability in the laser powder bed fusion process[J]. Clean Technologies and Recycling, 2022, 2(1): 32-46. doi: 10.3934/ctr.2022002
Additive manufacturing technologies such as laser powder bed fusion (LPBF) provide unprecedented abilities to manufacture complex 3-D parts and structures; however, the process produces leftover metal powder that is often unused, resulting in significant cost and disposal overhead. Towards this end, to ensure the ability to recycle the unused powder that is typically found within the build volume and in the overflow compartments of LPBF systems, a thorough chemical and structural analysis is required. Such an analysis will help in determining the extent of recyclability and reusability of the unused powder. For these purposes, characterization of recycled powder for Inconel 718 was performed in this work as a function of build-cycle to compare and contrast the evolution in physical and chemical properties such as particle size distribution, morphology, and composition. We note that the analysis was carried out for 21 build-cycles. Characterization of the particle size distribution and circularity was performed using traditional scanning electron microscopy methods and that information was later compared to measurements of surface profile using coherent scanning interferometry. Reusability was assessed using these results in conjunction with chemical analysis of the powder that was carried out using energy dispersive X-ray spectroscopy. For the extent of build-cycles considered in this work, it was seen that the physical and chemical changes in the recycled powder were will within accepted limits for powder usage and that with appropriate measures, recycled powder can be re-incorporated for subsequent builds without compromise in the powder quality. This study provides a pathway for greatly reducing powder consumption and thus cost during the LPBF process.

| [1] |
Wei C, Zhang Z, Cheng D, et al. (2020) An overview of laser-based multiple metallic material additive manufacturing: from macro-to micro-scales. Int J Extreme Manuf 3: 012003. https://doi.org/10.1088/2631-7990/abce04 doi: 10.1088/2631-7990/abce04

|
| [2] |
Gu DD, Meiners W, Wissenbach K, et al. (2012) Laser additive manufacturing of metallic components: materials, processes and mechanisms. Int Mater Rev 57: 133–164. https://doi.org/10.1179/1743280411Y.0000000014 doi: 10.1179/1743280411Y.0000000014
|
| [3] |
Hann BA (2016) Powder reuse and its effects on laser based powder fusion additive manufactured alloy 718. SAE Int J Aerosp 9: 209–214. https://doi.org/10.4271/2016-01-2071 doi: 10.4271/2016-01-2071
|
| [4] |
Slotwinski JA, Garboczi EJ, Stutzman PE, et al. (2014) Characterization of metal powders used for additive manufacturing. J Res Natl Inst Stan 119: 460. https://doi.org/10.6028/jres.119.018 doi: 10.6028/jres.119.018
|
| [5] |
Cordova L, Campos M, Tinga T (2019) Revealing the effects of powder reuse for selective laser melting by powder characterization. JOM 71: 1062–1072. https://doi.org/10.1007/s11837-018-3305-2 doi: 10.1007/s11837-018-3305-2
|
| [6] |
Ardila LC, Garciandia F, González-Díaz JB, et al. (2014) Effect of IN718 recycled powder reuse on properties of parts manufactured by means of selective laser melting. Phys Procedia 56: 99–107. https://doi.org/10.1016/j.phpro.2014.08.152 doi: 10.1016/j.phpro.2014.08.152
|
| [7] |
Paccou E, Mokhtari M, Keller C, et al. (2021) Investigations of powder reusing on microstructure and mechanical properties of Inconel 718 obtained by additive manufacturing. Mat Sci Eng A-Struct 828: 142113. https://doi.org/10.1016/j.msea.2021.142113 doi: 10.1016/j.msea.2021.142113
|
| [8] |
Emminghaus N, Hoff C, Hermsdorf J, et al. (2021) Residual oxygen content and powder recycling: Effects on surface roughness and porosity of additively manufactured Ti-6Al-4V. Addit Manuf 46: 102093. https://doi.org/10.1016/j.addma.2021.102093 doi: 10.1016/j.addma.2021.102093
|
| [9] |
Soundarapandiyan G, Johnston C, Khan RHU, et al. (2021) The effects of powder reuse on the mechanical response of electron beam additively manufactured Ti6Al4V parts. Addit Manuf 46: 102101. https://doi.org/10.1016/j.addma.2021.102101 doi: 10.1016/j.addma.2021.102101
|
| [10] |
Moghimian P, Poirié T, Habibnejad-Korayem M, et al. (2021) Metal powders in additive manufacturing: A review on reusability and recyclability of common titanium, nickel and aluminum alloys. Addit Manuf 43: 102017. https://doi.org/10.1016/j.addma.2021.102017 doi: 10.1016/j.addma.2021.102017
|
| [11] |
Fiegl T, Franke M, Raza A, et al. (2021) Effect of AlSi10Mg0.4 long-term reused powder in PBF-LB/M on the mechanical properties. Mater Design 212: 110176. https://doi.org/10.1016/j.matdes.2021.110176 doi: 10.1016/j.matdes.2021.110176
|
| [12] |
Gruber K, Smolina I, Kasprowicz M, et al. (2021) Evaluation of Inconel 718 metallic powder to optimize the reuse of powder and to improve the performance and sustainability of the laser powder bed fusion (LPBF) process. Materials 14: 1538. https://doi.org/10.3390/ma14061538 doi: 10.3390/ma14061538
|
| [13] |
Paccou E, Mokhtari M, Keller C, et al. (2021) Investigations of powder reusing on microstructure and mechanical properties of Inconel 718 obtained by additive manufacturing. Mat Sci Eng A-Struct 828: 142113. https://doi.org/10.1016/j.msea.2021.142113 doi: 10.1016/j.msea.2021.142113
|
| [14] |
Sendino S, Martinez S, Lamikiz A (2020) Characterization of IN718 recycling powder and its effect on LPBF manufactured parts. Procedia CIRP 94: 227–232. https://doi.org/10.1016/j.procir.2020.09.043 doi: 10.1016/j.procir.2020.09.043
|
| [15] |
Hilaire A, Andrieu E, Wu X (2019) High-temperature mechanical properties of alloy 718 produced by laser powder bed fusion with different processing parameters. Addit Manuf 26: 147–160. https://doi.org/10.1016/j.addma.2019.01.012 doi: 10.1016/j.addma.2019.01.012
|
| [16] |
Martin JH, Yahata BD, Hundley JM, et al. (2017) 3D printing of high-strength aluminium alloys. Nature 549: 365–369. https://doi.org/10.1038/nature23894 doi: 10.1038/nature23894
|
| [17] |
Jankovics D, Gohari H, Tayefeh M, et al. (2018) Developing topology optimization with additive manufacturing constraints in ANSYS®. IFAC-PapersOnLine 51: 1359–1364. https://doi.org/10.1016/j.ifacol.2018.08.340 doi: 10.1016/j.ifacol.2018.08.340
|
| [18] | Brown CU, Brown CU, Jacob G, et al. (2018) The effects of laser powder bed fusion process parameters on material hardness and density for nickel alloy 625, NIST Advanced Manufacturing Series. https://doi.org/10.6028/NIST.AMS.100-19 |
| [19] | Lane B, Mekhontsev S, Grantham S, et al. (2016) Design, developments, and results from the NIST additive manufacturing metrology testbed (AMMT). Solid Freeform Fabrication 2016: Proceedings of the 26th Annual International Solid Freeform Fabrication Symposium—An Additive Manufacturing Conference, 1145–1160. |
| [20] |
Bennett JL, Kafka OL, Liao H, et al. (2018) Cooling rate effect on tensile strength of laser deposited Inconel 718. Procedia Manuf 26: 912–919. https://doi.org/10.1016/j.promfg.2018.07.118 doi: 10.1016/j.promfg.2018.07.118
|
| [21] |
Hebert RJ (2016) Metallurgical aspects of powder bed metal additive manufacturing. J Mater Sci 51: 1165–1175. https://doi.org/10.1007/s10853-015-9479-x doi: 10.1007/s10853-015-9479-x
|
| [22] |
Savalani MM, Pizarro JM (2016) Effect of preheat and layer thickness on selective laser melting (SLM) of magnesium. Rapid Prototyp J 22: 115–122. https://doi.org/10.1108/RPJ-07-2013-0076 doi: 10.1108/RPJ-07-2013-0076
|
| [23] |
Averyanova M, Bertrand PH, Verquin B (2011) Studying the influence of initial powder characteristics on the properties of final parts manufactured by the selective laser melting technology. Virtual Phys Prototyp 6: 215–223. https://doi.org/10.1080/17452759.2011.594645 doi: 10.1080/17452759.2011.594645
|
| [24] |
Gomez C, Su R, Thompson A, et al. (2017) Optimization of surface measurement for metal additive manufacturing using coherence scanning interferometry. Opt Eng 56: 111714. https://doi.org/10.1117/1.OE.56.11.111714 doi: 10.1117/1.OE.56.11.111714
|
| [25] |
Townsend A, Senin N, Blunt L, et al. (2016) Surface texture metrology for metal additive manufacturing: a review. Precis Eng 46: 34–47. https://doi.org/10.1016/j.precisioneng.2016.06.001 doi: 10.1016/j.precisioneng.2016.06.001
|
| [26] |
DePond PJ, Guss G, Ly S, et al. (2018) In situ measurements of layer roughness during laser powder bed fusion additive manufacturing using low coherence scanning interferometry. Mater Design 154: 347–359. https://doi.org/10.1016/j.matdes.2018.05.050 doi: 10.1016/j.matdes.2018.05.050
|
| [27] | Material Data Sheet EOS NickelAlloy IN718, Material Data Sheet Technical Data. EOS, 2022. Available from: https://www.eos.info/03_system-related-assets/material-related-contents/metal-materials-and-examples/metal-material-datasheet/nickelalloy-inconel/material_datasheet_eos_nickelalloy_in718_en.pdf. |
| [28] | Ferreira T, Rasband W (2012) ImageJ User Guide. Available from: https://imagej.nih.gov/ij/docs/guide/. |






Figures(7) / Tables(3)
Susana J. Castillo, Anna Hayes, Greg Colvin, Barrett G. Potter, Rongguang Liang, Krishna Muralidharan. Characterization of recycled Inconel 718 metal powder for assessing its reusability in the laser powder bed fusion process[J]. Clean Technologies and Recycling, 2022, 2(1): 32-46. doi: 10.3934/ctr.2022002







 DownLoad:
DownLoad: