Cardiac mitochondria are intracellular organelles that play an important role in energy metabolism and cellular calcium regulation. In particular, they influence the excitation-contraction cycle of the heart cell. A large number of mathematical models have been proposed to better understand the mitochondrial dynamics, but they generally show a high level of complexity, and their parameters are very hard to fit to experimental data.
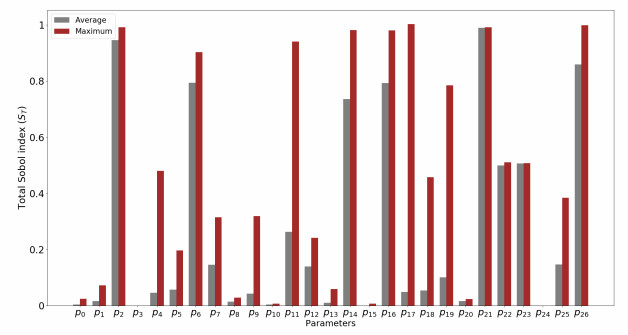
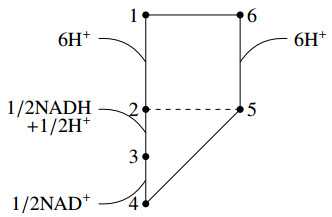
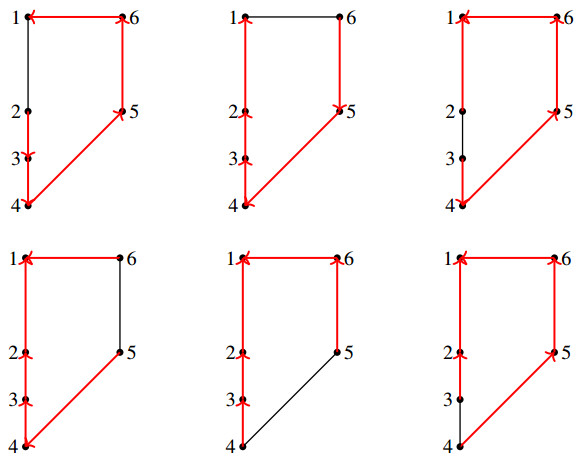
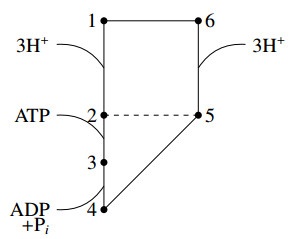
We derived a model based on historical free energy-transduction principles, and results from the literature. We proposed simple expressions that allow to reduce the number of parameters to a minimum with respect to the mitochondrial behavior of interest for us. The resulting model has thirty-two parameters, which are reduced to twenty-three after a global sensitivity analysis of its expressions based on Sobol indices.
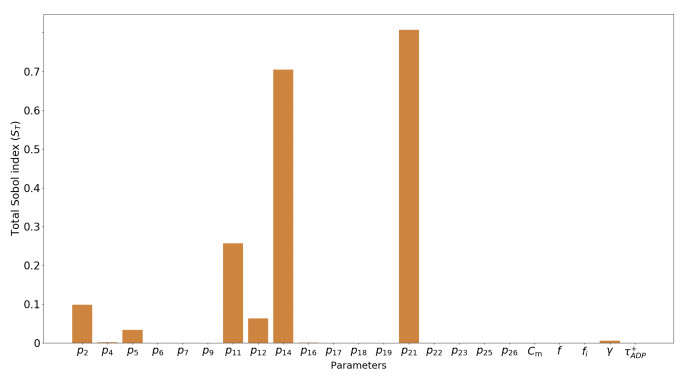
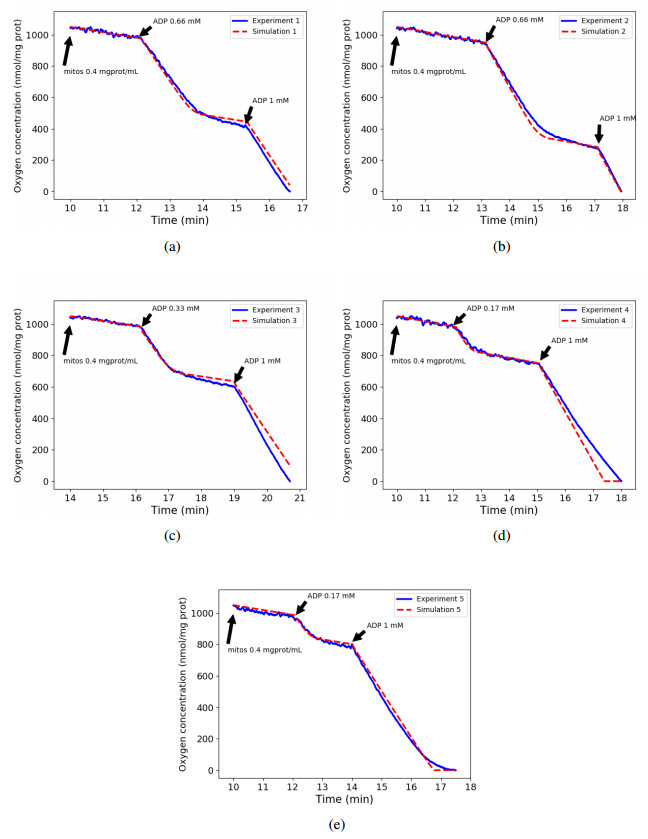
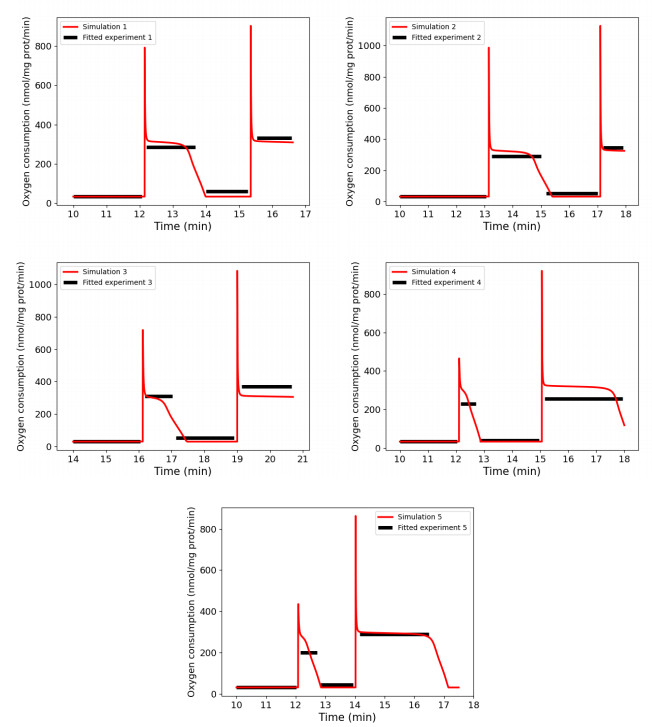
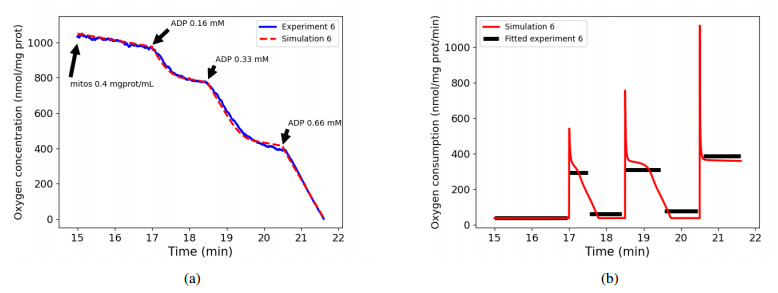
We calibrated our model to experimental data that consists of measurements of mitochondrial respiration rates controlled by external ADP additions. A sensitivity analysis of the respiration rates showed that only seven parameters can be identified using these observations. We calibrated them using a genetic algorithm, with five experimental data sets. At last, we used the calibration results to verify the ability of the model to accurately predict the values of a sixth dataset.
Results show that our model is able to reproduce both respiration rates of mitochondria and transitions between those states, with very low variability of the parameters between each experiment. The same methodology may apply to recover all the parameters of the model, if corresponding experimental data were available.
Citation: Bachar Tarraf, Emmanuel Suraniti, Camille Colin, Stéphane Arbault, Philippe Diolez, Michael Leguèbe, Yves Coudière. A simple model of cardiac mitochondrial respiration with experimental validation[J]. Mathematical Biosciences and Engineering, 2021, 18(5): 5758-5789. doi: 10.3934/mbe.2021291
Cardiac mitochondria are intracellular organelles that play an important role in energy metabolism and cellular calcium regulation. In particular, they influence the excitation-contraction cycle of the heart cell. A large number of mathematical models have been proposed to better understand the mitochondrial dynamics, but they generally show a high level of complexity, and their parameters are very hard to fit to experimental data.
We derived a model based on historical free energy-transduction principles, and results from the literature. We proposed simple expressions that allow to reduce the number of parameters to a minimum with respect to the mitochondrial behavior of interest for us. The resulting model has thirty-two parameters, which are reduced to twenty-three after a global sensitivity analysis of its expressions based on Sobol indices.
We calibrated our model to experimental data that consists of measurements of mitochondrial respiration rates controlled by external ADP additions. A sensitivity analysis of the respiration rates showed that only seven parameters can be identified using these observations. We calibrated them using a genetic algorithm, with five experimental data sets. At last, we used the calibration results to verify the ability of the model to accurately predict the values of a sixth dataset.
Results show that our model is able to reproduce both respiration rates of mitochondria and transitions between those states, with very low variability of the parameters between each experiment. The same methodology may apply to recover all the parameters of the model, if corresponding experimental data were available.

| [1] | P. Diolez, V. Deschodt-Arsac, G. Raffard, C. Simon, P. D. Santos, E. Thiaudière, et al., Modular regulation analysis of heart contraction: application to in situ demonstration of a direct mitochondrial activation by calcium in beating heart, Am. J. Physiol.-Reg., I., 293 (2007), R13–R19. |
| [2] | M. Luo, M. E. Anderson, Mechanisms of altered Ca2+ handling in heart failure, Circ. Res., 113 (2013), 690–708. |
| [3] | A. Takeuchi, S. Matsuoka, Integration of mitochondrial energetics in heart with mathematical modelling, J. Physiol.-London, 598 (2020), 1443–1457. |
| [4] |
G. Magnus, J. Keizer, Minimal model of $\beta$-cell mitochondrial Ca$^{2+}$ handling, Am. J. Physiol.-Cell Ph., 273 (1997), C717–C733. doi: 10.1152/ajpcell.1997.273.2.C717

|
| [5] |
R. Bertram, M. G. Pedersen, D. S. Luciani, A. Sherman, A simplified model for mitochondrial ATP production, J. Theor. Biol., 243 (2006), 575–586. doi: 10.1016/j.jtbi.2006.07.019
|
| [6] | C. C. Mitchell, D. G. Schaeffer, A two-current model for the dynamics of cardiac membrane, B. Math. Biol., 65 (2003), 767–793. |
| [7] | K. H. ten Tusscher, A. V. Panfilov, Alternans and spiral breakup in a human ventricular tissue model, Am. J. Physiol. Heart C., 291 (2006), H1088–1100. |
| [8] | T. L. Hill, Free energy transduction in biology: the steady-state kinetic and thermodynamic formalism, Academic Press, 1977. |
| [9] |
D. Pietrobon, S. R. Caplan, Flow-force relationships for a six-state proton pump model: intrinsic uncoupling, kinetic equivalence of input and output forces, and domain of approximate linearity, Biochemistry-US, 24 (1985), 5764–5776. doi: 10.1021/bi00342a012
|
| [10] | M.-H. T. Nguyen, M. S. Jafri, Mitochondrial calcium signaling and energy metabolism, Ann. N.Y. Acad. Sci., 1047 (2005), 127–137. |
| [11] | M.-H. T. Nguyen, S. J. Dudycha, M. S. Jafri, Effect of Ca2+ on cardiac mitochondrial energy production is modulated by Na+ and H+ dynamics, Am. J. Physiol.-Cell Ph., 292 (2007), C2004–C2020. |
| [12] |
S. Cortassa, M. A. Aon, E. Marbân, R. L. Winslow, B. O'Rourke, An integrated model of cardiac mitochondrial energy metabolism and calcium dynamics, Biophys. J., 84 (2003), 2734–2755. doi: 10.1016/S0006-3495(03)75079-6
|
| [13] | S. Cortassa, B. O'Rourke, R. L. Winslow, M. A. Aon, Control and regulation of mitochondrial energetics in an integrated model of cardiomyocyte function, Biophys. J., 96 (2009), 2466–2478. |
| [14] |
L. D. Gauthier, J. L. Greenstein, S. Cortassa, B. O'Rourke, R. L. Winslow, A computational model of reactive oxygen species and redox balance in cardiac mitochondria, Biophys. J., 105 (2013), 1045–1056. doi: 10.1016/j.bpj.2013.07.006
|
| [15] | G. Magnus, J. Keizer, Model of $\beta$-cell mitochondrial calcium handling and electrical activity. I. Cytoplasmic variables, Am. J. Physiol.-Cell Ph., 274 (1998), C1158–C1173. |
| [16] | M. D. Brand, The stoichiometry of the exchange catalysed by the mitochondrial calcium/sodium antiporter, Biochem. J., 229 (1985), 161–166. |
| [17] | T. Matsuda, K. Takuma, A. Baba, Na+-Ca2+ exchanger: physiology and pharmacology, Jpn. J. Pharmacol., 74 (1997), 1–20. |
| [18] | T. E. Gunter, D. R. Pfeiffer, Mechanisms by which mitochondria transport calcium, Am. J. Physiol.-Cell Ph., 258 (1990), C755–C786. |
| [19] |
D. E. Wingrove, T. Gunter, Kinetics of mitochondrial calcium transport. I. Characteristics of the sodium-independent calcium efflux mechanism of liver mitochondria, J. Biol. Chem., 261 (1986), 15159–15165. doi: 10.1016/S0021-9258(18)66846-2
|
| [20] | G. C. Brown, The leaks and slips of bioenergetic membranes, FASEB J., 6 (1992), 2961–2965. |
| [21] | M. D. Brand, L.-F. Chien, P. Diolez, Experimental discrimination between proton leak and redox slip during mitochondrial electron transport, Biochem. J., 297 (Pt 1) (1994), 27–29. |
| [22] | G. Krishnamoorthy, P. C. Hinkle, Non-ohmic proton conductance of mitochondria and liposomes, Biochemistry-US, 23 (1984), 1640–1645. |
| [23] | C. Colin, Développement de nouvelles méthodes de microscopie et d'électrochimie pour des analyses multi-paramétriques de mitochondries individuelles cardiaques (in French), Theses, Université de Bordeaux, 2020, Available from: https://tel.archives-ouvertes.fr/tel-03116368. |
| [24] | Carlos M. Palmeira, António J. Moreno (eds.), Mitochondrial Bioenergetics, vol. 1782 of Methods in Molecular Biology, Humana Press, 2008. |
| [25] |
B. Chance, W. G.R., Respiratory enzymes in oxidative phosphorylation: III. The steady state, J. Biol. Chem., 217 (1955), 409–427. doi: 10.1016/S0021-9258(19)57191-5
|
| [26] | A. Saltelli, M. Ratto, T. Andres, F. Campolongo, J. Cariboni, D. Gatelli, et al., Global sensitivity analysis: the primer, John Wiley & Sons, 2008. |
| [27] |
J. Herman, W. Usher, SALib: An open-source Python library for Sensitivity Analysis, J. Open Source Softw., 2 (2017), 97. doi: 10.21105/joss.00097
|
| [28] |
F. Wu, F. Yang, K. Vinnakota, D. Beard, Computer modeling of mitochondrial tricarboxylic acid cycle, oxidative phosphorylation, metabolite transport, and electrophysiology, J. Biol. Chem., 282 (2007), 24525–24537. doi: 10.1074/jbc.M701024200
|
 mbe-18-05-291-s001.zip mbe-18-05-291-s001.zip |
 |

Figures(14) / Tables(9)

Bachar Tarraf, Emmanuel Suraniti, Camille Colin, Stéphane Arbault, Philippe Diolez, Michael Leguèbe, Yves Coudière. A simple model of cardiac mitochondrial respiration with experimental validation[J]. Mathematical Biosciences and Engineering, 2021, 18(5): 5758-5789. doi: 10.3934/mbe.2021291







 DownLoad:
DownLoad: