Finance has an important influence on technological innovation (TI). There are several stages of, as well as various financial constraints on, TI. In this article, we divide TI into four stages: the development, growth, maturity, and decline stages. Concurrently, we classify TI funding sources into five types: enterprise funds, government funds, venture capital funds, loans from financial institutions, and capital market funds. Based on the analysis of the stages and financing constraints of TI, this paper constructs a state-space model to study the effects of various funding sources on TI in Hebei Province, China, from 2005 to 2018. The results show a long-term equilibrium relationship between finance and TI, whereby different financial methods have different effects on each stage of TI. Enterprise funds play a primary role in the development, growth, and maturity stages. Government funds play a prominent role in the development and growth stages. Capital market funds and loans from financial institutions only play a role in the maturity period. The role of capital market funds was positive, while that of loans from financial institutions was negative. The effect of venture capital was not noticeable at any stage. Finally, we give our conclusions and put forward some countermeasures and suggestions to promote TI in Hebei Province.
Citation: Mengxin Wang, Ran Gu, Meng Wang, Junru Zhang. Research on the impact of finance on promoting technological innovation based on the state-space model[J]. Green Finance, 2021, 3(2): 119-137. doi: 10.3934/GF.2021007
Finance has an important influence on technological innovation (TI). There are several stages of, as well as various financial constraints on, TI. In this article, we divide TI into four stages: the development, growth, maturity, and decline stages. Concurrently, we classify TI funding sources into five types: enterprise funds, government funds, venture capital funds, loans from financial institutions, and capital market funds. Based on the analysis of the stages and financing constraints of TI, this paper constructs a state-space model to study the effects of various funding sources on TI in Hebei Province, China, from 2005 to 2018. The results show a long-term equilibrium relationship between finance and TI, whereby different financial methods have different effects on each stage of TI. Enterprise funds play a primary role in the development, growth, and maturity stages. Government funds play a prominent role in the development and growth stages. Capital market funds and loans from financial institutions only play a role in the maturity period. The role of capital market funds was positive, while that of loans from financial institutions was negative. The effect of venture capital was not noticeable at any stage. Finally, we give our conclusions and put forward some countermeasures and suggestions to promote TI in Hebei Province.

| [1] |
Amore MD, Schneider C, Aldokas A (2013) Credit supply and corporate innovation. J Financ Econ 109: 835-855. doi: 10.1016/j.jfineco.2013.04.006

|
| [2] |
Andrew T (2007) The role of finance and corporate governance in national systems of innovation. Organ Stud 28: 1461-1481. doi: 10.1177/0170840607075676
|
| [3] | Atanassov J, Nanda VK, Seru A (2007) Finance and innovation: The case of publicly traded firms. SSRN Electron J 15: 89-92. |
| [4] |
Audretsch DB, Lehmann EE, Paleari S, et al. (2016) Entrepreneurial finance and technology transfer. J Technol Transfer 41: 1-9. doi: 10.1007/s10961-014-9381-8
|
| [5] |
Bejaković P (2002) The financing of research and development. Oxf Rev Econ Policy 18: 35-51. doi: 10.1093/oxrep/18.1.35
|
| [6] | China Venture Capital Research Institute (2010-2018) China Venture Capital Yearbook, Beijing: China Development Press. |
| [7] | Chinese Academy of Science and Technology for Development (2005-2009) Venture Capital Development in China, Beijing: Economic & Management Publishing House. |
| [8] |
Chowdhury RH, Maung M (2012) Financial market development and the effectiveness of R & D investment: Evidence from developed and emerging countries. Res Int Bu Financ 26: 258-272. doi: 10.1016/j.ribaf.2011.12.003
|
| [9] |
Dosi G (1990) Finance, innovation and industrial change. J Econ Behav Organ 13: 299-319. doi: 10.1016/0167-2681(90)90003-V
|
| [10] | Dushnitsky G, Lenox M (2005) When do incumbents learn from entrepreneurial ventures? Res Policy 34: 615-639. |
| [11] | Economics V (1983) Regional Patterns of Venture Capital Investment. Prepared for US Small Business Administration. Washington, DC: Small Business Administration. |
| [12] | Gao H (2017) Research on the Effectiveness of Supporting Scientific and Technological Innovation by Science and Technology Finance in Henan Province—An empirical Test based on panel Model. Financ Theory Pract, 65-70. |
| [13] |
Giudici G, Paleari S (2000) The provision of finance to innovation: A survey conducted among Italian technology-based small firms. Small Bus Econ 14: 37-53. doi: 10.1023/A:1008187416389
|
| [14] |
Greenwood J, Jovanovic B (1990) Financial Development, Growth, and the Distribution of Income. J Political Economy 98: 1076-1107. doi: 10.1086/261720
|
| [15] | Hebei Provincial People's Government (2005-2019) Hebei Economic Yearbook, Beijing: China Statistics Press. |
| [16] |
King RG, Levine R (1993) Finance, entrepreneurship and growth: Theory and evidence. J Monetary Econ 32: 513-542. doi: 10.1016/0304-3932(93)90028-E
|
| [17] |
Law SH, Lee WC, Singh N (2018) Revisiting the finance-innovation nexus: Evidence from a non-linear approach. J Innov Knowl 3: 143-153. doi: 10.1016/j.jik.2017.02.001
|
| [18] |
Legrand DP, Pommet S (2010) Venture capital syndication and the financing of innovation: Financial versus expertise motives. Econ Lett 106: 75-77. doi: 10.1016/j.econlet.2009.10.004
|
| [19] |
Lerner J, Nanda R (2020) Venture capital's role in financing innovation: What we know and how much we still need to learn. J Econ Perspect 34: 237-261. doi: 10.1257/jep.34.3.237
|
| [20] |
Levine R (1991) Stock Markets, Growth, and Tax Policy. J Financ 46: 1445-1465. doi: 10.1111/j.1540-6261.1991.tb04625.x
|
| [21] | Liang L, Ma R, Tian Y (2009) R & D Financing Source and Technical Innovation—Empirical Research on China's Large and Medium-Size Industry Firms. Sci Sci Manage S T 7: 89-93. |
| [22] |
Liao G, Drakeford BM (2019) An analysis of financial support, technological progress and energy efficiency:evidence from China. Green Financ 1: 174-187. doi: 10.3934/GF.2019.2.174
|
| [23] | Markard J (2020) The life cycle of technological innovation systems. Technol Forecast Soc Change 153: 119407. |
| [24] |
Nanda R, Rhodes-Kropf M (2016) Financing risk and innovation. Manage Sci 63: 901-918. doi: 10.1287/mnsc.2015.2350
|
| [25] | National Bureau of Statistics, Ministry of Science and Technology (2005-2019) China Statistics Yearbook on Science and Technology, Beijing: China Statistics Press. |
| [26] | Perez C (2013) Finance and technical change: a neo-schumpeterian perspective. Financ Market Res: 76-91. |
| [27] | Pu Z, Tian Q, Jiangpeng Z (2017) Research on the Influence of Technology Finance on Technology Innovation. 2017 3rd International Conference on Innovation Development of E-commerce and Logistics (ICIDEL 2017), 190-194. |
| [28] |
Rin MD, Nicodano G, Sembenelli A (2006) Public policy and the creation of active venture capital markets. J Public Econ 90: 1699-1723. doi: 10.1016/j.jpubeco.2005.09.013
|
| [29] | Rosenberg N (2009) Studies on science and the innovation process. World Sci, 428. |
| [30] |
Saint Paul G (1992) Technological Choice, Financial Markets and Economic Development. Eur Econ Rev 36: 763-781. doi: 10.1016/0014-2921(92)90056-3
|
| [31] |
Sasidharan S, Jijo Lukose PJ, Komera S (2015) Financing constraints and investments in R & D: Evidence from Indian manufacturing firms. Quart Revi Econ Financ 55: 28-39. doi: 10.1016/j.qref.2014.07.002
|
| [32] | Schumpeter JA (1934) The theory of economic development; an inquiry into profits, capital, credit, interest, and the business cycle, Cambridge, Mass.: Harvard University Press. |
| [33] | Science and Technology Department of Hebei Province, Hebei Provincial Statistics Bureau (2005-2019) Hebei Statistics Yearbook on Science and Technology, Shijiazhuang: Hebei Science & Technology Press. |
| [34] | Siming L, Mengxin W, Yong T (2019) Stabilizing inflation expectations in China: Does economic policy uncertainty matter? Green Financ 1: 429-441. |
| [35] |
Song M, Ai H, Li X (2015) Political connections, financing constraints, and the optimization of innovation efficiency among China's private enterprises. Technol Forecast Soc Change 92: 290-299. doi: 10.1016/j.techfore.2014.10.003
|
| [36] |
Kaplan SN, Strömberg P (2003) Financial contracting theory meets the real world: An empirical analysis of venture capital contracts. Rev Econ Stud 70: 281-315. doi: 10.1111/1467-937X.00245
|
| [37] |
Stulz RM (2000) Financial structure, corporate finance and economic growth. Int Rev Financ 1: 11-38. doi: 10.1111/1468-2443.00003
|
| [38] |
Timmons JA, Bygrave WD (1986) Venture capital, s role in financing innovation for economic growth. J Bus Venturing 1: 161-176. doi: 10.1016/0883-9026(86)90012-1
|
| [39] |
Vernon R (1966) International investment and international trade in the product cycle. Quart J Econ 80: 190-207. doi: 10.2307/1880689
|
| [40] | Yu L (2013) Empirical Study on Regional Finance and S & T Innovation. Sci Sci Manage S T 34: 88-97. |
| [41] |
Zhang C, Mao D, Wang M (2020) Role of Venture Capital in Enterprise Innovation Under Psychological Capital and Heterogeneity of Entrepreneur Capital. Front Psychol 11: 1704-1704. doi: 10.3389/fpsyg.2020.01704
|
| [42] |
Zhang L, Zhang S, Guo Y (2019) The effects of equity financing and debt financing on technological innovation: Evidence from developed countries. Baltic J Manage 14: 698-715. doi: 10.1108/BJM-01-2019-0011
|
| [43] | Zhang Y, Zhao L (2015) The effect of sci-tech finance investment on sci-tech innovation in Chowdhury: An empirical research based on the static and dynamic panel date model. Stud Sci Sci 33: 177-184. |
| [44] | Zheng Y, LI Z (2015) Research on the Effectiveness of China's Financing Service for Technology Innovation. China Soft Sci, 127-136. |
| [45] | Zhu E, Zhang Q, Sun L (2021) Enterprise financing mode and technological innovation behavior selection: An empirical analysis based on the data of the World Bank's survey of Chinese private enterprises. Discrete Dyn Nat Soc 2021: 8833979. |
| [46] | Zhu Y, Chen L (2008) Research on the development of high-tech industries' finance. J Guizhou Norm Univ, 53-58. |
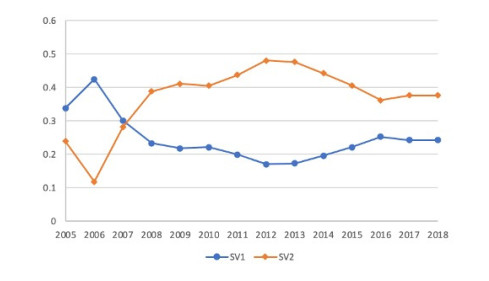
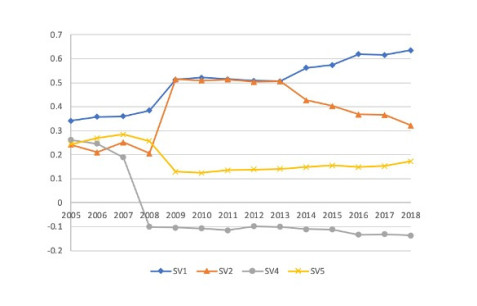
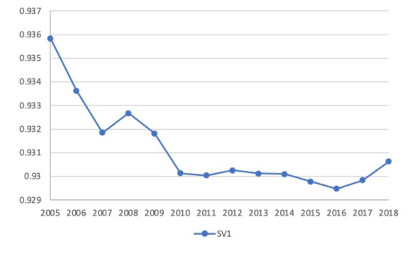
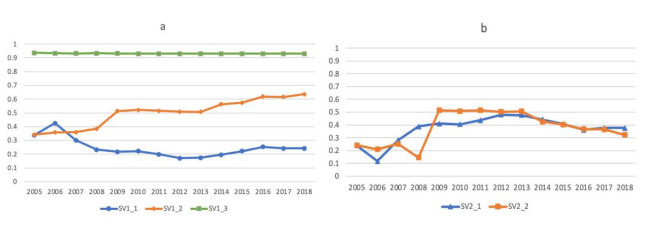
Figures(4) / Tables(7)

Mengxin Wang, Ran Gu, Meng Wang, Junru Zhang. Research on the impact of finance on promoting technological innovation based on the state-space model[J]. Green Finance, 2021, 3(2): 119-137. doi: 10.3934/GF.2021007







 DownLoad:
DownLoad: