Citation: Keiji Sakakibara, Takashi Kanamura. Risk of temperature differences in geothermal wells and generation strategies of geothermal power[J]. Green Finance, 2020, 2(4): 424-436. doi: 10.3934/GF.2020023

| [1] |
Alexander GB, Heston WM, Iler RK (1954) The solubility of amorphous silica in water. J Phys Chem 58: 453-455. doi: 10.1021/j150516a002

|
| [2] |
Anderson A, Rezaie B (2019) Geothermal technology: Trends and potential role in a sustainable future. Appl Energ 248: 18-34. doi: 10.1016/j.apenergy.2019.04.102
|
| [3] | Bilqist RA, Dachyar M, Farizal (2018) The valuation of geothermal power projects in Indonesia using real options valuation, In: MATEC Web of Conferences, EDP Sciences, 248: 03004. |
| [4] |
Chen S, Zhang Q, Tang Y, et al. (2019) Investment strategy for shallow geothermal resource based on real option model. Energ Procedia 158: 6118-6125. doi: 10.1016/j.egypro.2019.01.500
|
| [5] |
Compernolle T, Welkenhuysen K, Petitclerc E, et al. (2019) The impact of policy measures on profitability and risk in geothermal energy investments. Energ Econ 84: 104524. doi: 10.1016/j.eneco.2019.104524
|
| [6] | Thermal Nuclear Power Technology Association (2018) Current Status and Trends in Geothermal Power Generation 2017. 3: 43-44 (in Japanese). |
| [7] |
Feili HR, Akar N, Lotfizadeh H, et al. (2013) Risk analysis of geothermal power plants using failure modes and effects analysis (FMEA) technique. Energ Convers Manage 72: 69-76. doi: 10.1016/j.enconman.2012.10.027
|
| [8] | Founier RO, Rowe JJ (1977) The solubility of amorphous silica in water at high temperatures and high pressures. Am Mineral 62: 1052-1056. |
| [9] | Gazheli A, van den Bergh J (2018) Real options analysis of investment in solar vs. wind energy: Diversification strategies under uncertain prices and costs. Renew Sust Energ Rev 82: 2693-2704. |
| [10] | Goto K (1955) Studies on the State of Silicate in Water (Part 2) Solubility in Amorphous Calculations. Jpn J Chem 76: 1364-1366 (in Japanese). |
| [11] | Hirowatari K (1990) Experimental study on scale prevention method using exhausted gases from geothermal power station. J Geotherm Res Soc Japan 12: 347-362 (in Japanese). |
| [12] | Hosoi M, Imai H (1982) Study on precipitation and prevention of the silica scale from the geothermal hot water. J Geotherm Res Soc Japan 4: 127-142 (in Japanese). |
| [13] | Huttrer GW (2020) Geothermal Power Generation in the World 2015-2020 Update Report. Proceedings World Geothermal Congress 2020 Reykjavik, Iceland. |
| [14] | IEA Geothermal (2020) Annual Report 2019. Available from: https://drive.google.com/file/d/1hqz5BB391z_LcaeVERQ_YU5zJbhm-2Ok/view. |
| [15] | Itoi R, Maekawa H, Fukuda M, et al. (1986) Study on decrease of reservoir permeability due to deposition of silica dissolved in reinjection water. J Geotherm Res Soc Japan 8: 229-241 (in Japanese). |
| [16] | Itoi R, Mekawa H, Fukuda M, et al. (1987) Decrease in reservoir permeability due to deposition of silica dissolved in geothermal injection water numerical simulation in one dimensional flow system. J Geother Res Soc Japan 9: 285-306 (in Japanese). |
| [17] |
Karadas M, Celik HM, Serpen U, et al. (2015) Multiple regression analysis of performance parameters of a binary cycle geothermal power plant. Geothermics 54: 68-75. doi: 10.1016/j.geothermics.2014.11.003
|
| [18] | Kimbara K (1999) Geothermal resources survey in the volcanic fields of Japan: a review. Earth Sci (Chikyu Kagaku) 53: 325-399 (in Japanese). |
| [19] |
Kitzing L, Juul N, Drud M, et al. (2017) A real options approach to analyse wind energy investments under different support schemes. Appl Energ 188: 83-96. doi: 10.1016/j.apenergy.2016.11.104
|
| [20] | Knaut A, Madlener R, Rosen C, et al. (2012) Effects of temperature uncertainty on the valuation of geothermal projects: a real options approach. FCN Working Paper 11. |
| [21] |
Kojima Y, Kawanabe A, Yasue T, et al. (1993) Synthesis of amorphous calcium carbonate and its crystallization. J Ceramic Soc Japan 101: 1145-1152 (in Japanese). doi: 10.2109/jcersj.101.1145
|
| [22] |
Loncar D, Milovanovic I, Rakic B, et al. (2017) Compound real options valuation of renewable energy projects: The case of a wind farm in Serbia. Renew Sust Energ Rev 75: 354-367. doi: 10.1016/j.rser.2016.11.001
|
| [23] | Ministry of the Environment (2010) Study on the Method of Reviewing Environmental Impact Related to Geothermal Power Generation (in Japanese). Available from: https://www.env.go.jp/nature/geothermal_power/conf/h2304/ref02.pdf. |
| [24] |
Mo J, Schleich J, Fan Y (2018) Getting ready for future carbon abatement under uncertainty—Key factors driving investment with policy implications. Energ Econ 70: 453-464. doi: 10.1016/j.eneco.2018.01.026
|
| [25] | New Energy and Industrial Technology Development Organization (2014) Renewable Energy Technology White Paper (in Japanese). Available from: https://www.nedo.go.jp/content/100544822.pdf. |
| [26] |
Ritzenhofen I, Spinler S (2016) Optimal design of feed-in-tariffs to stimulate renewable energy investments under regulatory uncertainty—A real options analysis. Energ Econ 53: 76-89. doi: 10.1016/j.eneco.2014.12.008
|
| [27] | Sasaki M (2018) Classification of water types of acid hot-spring waters in Japan. J Geotherm Res Soc Japan 40: 235-243. |
| [28] | Sato K, Odanaka K, Shakunaga N, et al. (1996) How to inject water into the geothermal fluid transport pipe. Japan Metals and Chemicals Co., Ltd. Jan30th, 1996, Patent Number: 1996-028432 (in Japanese). |
| [29] | SPATECH Shinshu (2015) (in Japanese). Available from: http://spatec-shinshu.jp/c3b.html. Access on: Dec 3, 2020. |
| [30] | Snyder DM, Beckers KF, Young KR, et al. (2017) Analysis of geothermal reservoir and well operational conditions using monthly production reports from Nevada and California. GRC Trans 41: 2844-2856. |
| [31] |
Shiozaki A (2019) Geothermal power generation development in Japan. Japan Soc Eng Geol 60: 120-124 (In Japanese). doi: 10.5110/jjseg.60.120
|
| [32] | Yu S, Li Z, Wei Y, et al. (2019) A real option model for geothermal heating investment decision making: Considering carbon trading and resource taxes. Energy 189: 116252. |
| [33] | Zhang M, Liu L, Wang Q, et al. (2020) Valuing investment decisions of renewable energy projects considering changing volatility. Energ Econ 92: 104954. |
 GF-02-04-023-s001.pdf GF-02-04-023-s001.pdf |
 |
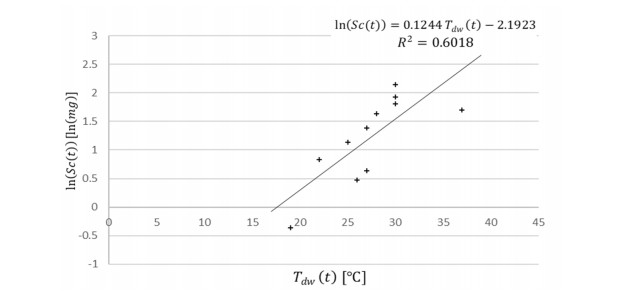
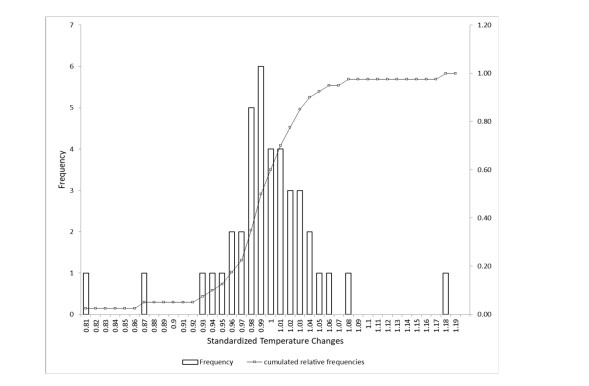
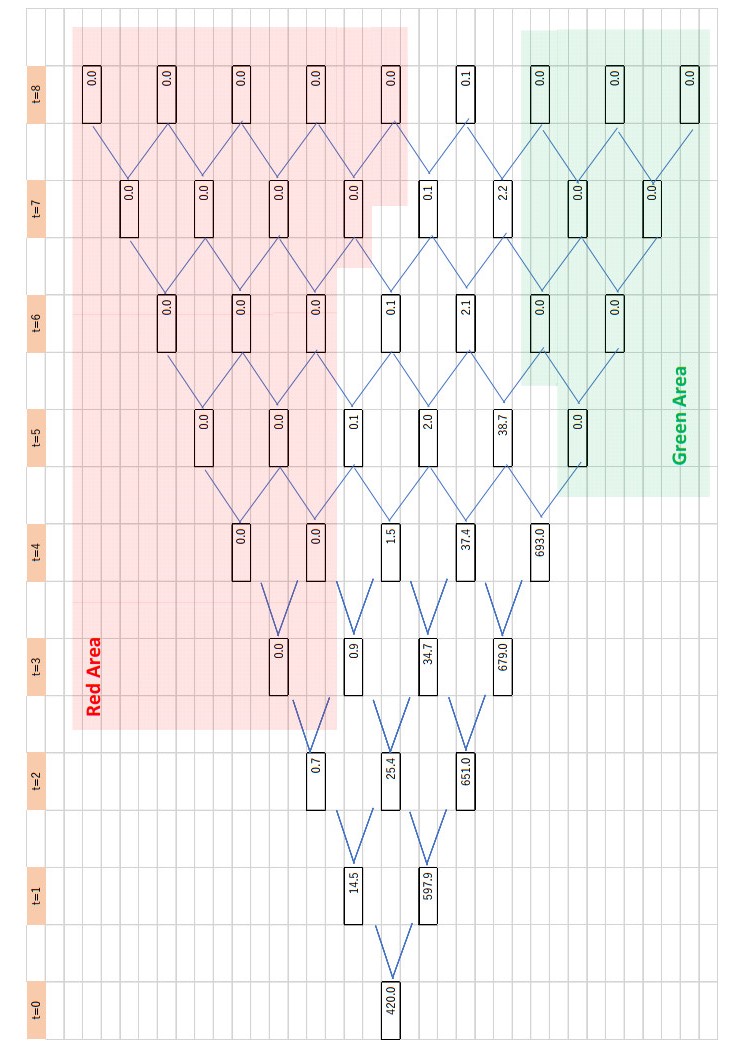
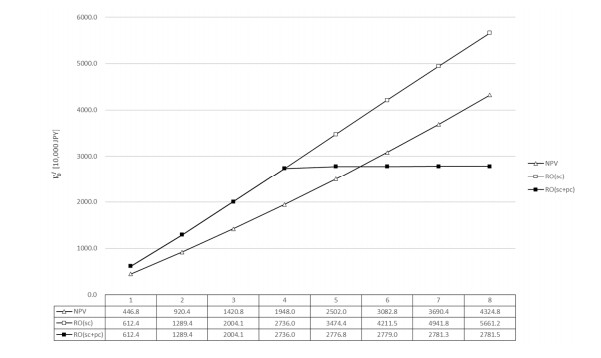
Figures(4) / Tables(2)

Keiji Sakakibara, Takashi Kanamura. Risk of temperature differences in geothermal wells and generation strategies of geothermal power[J]. Green Finance, 2020, 2(4): 424-436. doi: 10.3934/GF.2020023







 DownLoad:
DownLoad: