Citation: Gashaw Beyene, Gamachis Sakata, Teshome Senbeta, Belayneh Mesfin. Effect of core size/shape on the plasmonic response of spherical ZnO@Au core-shell nanostructures embedded in a passive host-matrices of MgF2[J]. AIMS Materials Science, 2020, 7(6): 705-719. doi: 10.3934/matersci.2020.6.705

| [1] | Ismail MM, Cao WQ, Humadi MD (2016) Synthesis and optical properties of Au/ZnO core-shell nanorods and their photocatalytic activities. Optik 127: 4307-4311. |
| [2] | Brinson BE, Lassiter JB, Levin CS, et al. (2008) Nanoshells made easy: Improving Au layer growth on nanoparticle surfaces. Langmuir 24: 14166-14171. |
| [3] | Azizi S, Mohamad R, Rahim RA, et al. (2016) ZnO-Ag core shell nanocomposite formed by green method using essential oil of wild ginger and their bactericidal and cytotoxic effects. Appl Surf Sci 384: 517-524. |
| [4] | Bartosewicz B, Michalska-Domańska M, Liszewska M, et al. (2017) Synthesis and characterization of noble metal-titania core-shell nanostructures with tunable shell thickness. Beilstein J Nanotechno 8: 2083-2093. |
| [5] | Fan CZ, Wang JQ, Cheng YG, et al. (2013) Electric field distribution around the chain of composite nanoparticles in ferrofluids. Chinese Phys B 22: 1-6. |
| [6] | Sadollahkhani A, Kazeminezhad I, Lu J, et al. (2014) Synthesis, structural characterization and photocatalytic application of ZnO@ZnS core-shell nanoparticles. RSC Adv 4: 36940-36950. |
| [7] | Kassahun GB (2019) High tunability of size dependent optical properties of ZnO@M@Au (M = SiO2, TiO2, In2O3) core/spacer/shell nanostructure. Adv Nano Res 2: 1-13. |
| [8] | Encina ER, Prez MA, Coronado EA (2013) Synthesis of Ag@ZnO core-shell hybrid nanostructures: an optical approach to reveal the growth mechanism. J Nanopart Res 15: 1688. |
| [9] | Derkachova A, Kolwas K, Demchenko I (2016) Dielectric function for gold in plasmonics applications: Size dependence of plasmon resonance frequencies and damping rates for nanospheres. Plasmonics 11: 941-951. |
| [10] | Wang B, Zhu X, Li S, et al. (2018) Ag@SiO2 core-shell nanoparticles embedded in a TiO2 mesoporous layer substantially improve the performance of perovskite solar cells. Nanomaterials 8: 701. |
| [11] | Bai Y, Butburee T, Yu H, et al. (2015) Controllable synthesis of concave cubic gold core-shell nanoparticles for plasmon-enhanced photon harvesting. J Colloid Interf Sci 449: 246-251. |
| [12] | Daneshfar N, Bazyari K (2014) Optical and spectral tunability of multilayer spherical and cylindrical nanoshells. Appl Phys A-Mater 116: 611-620. |
| [13] | Elyahb AK, Elise C, Yongmei W, et al. (2017) Synthesis and properties of magnetic optical core-shell nanoparticles. RSC Adv 7: 17137. |
| [14] | Alzahrani E (2017) Photodegradation of binary AZO dyes using core-shell Fe3O4/SiO2/TiO2 nanospheres. Am J Anal Chem 8: 95115. |
| [15] | Shao X, Li B, Zhang B, et al. (2016) Au@ZnO core-shell nanostructures with plasmon-induced visible-light photocatalytic and photoelectrochemical properties. Inorg Chem Front 3: 934-943. |
| [16] | Wang H, Zhou L, Chu X, et al. (2018) Band alignment of BiOCl/ZnO core shell nanosheets by X-ray photoelectron spectroscopy measurements. Ferroelectrics 531: 31-37. |
| [17] | Li J, Cushing SK, Bright J, et al. (2013) Ag@Cu2O core-shell nanoparticles as visible-light plasmonic photocatalysts. ACS Catal 3: 47-51. |
| [18] | He L, Liu Y, Liu J, et al. (2013) Core-shell noble-metal@metal-organic-framework nanoparticles with highly selective sensing property. Angew Chem 125: 3829-3833. |
| [19] | Lee S, Lee J, Nam K, et al. (2016) Application of Ni-oxide@TiO2 core-shell structures to photocatalytic mixed dye degradation, CO oxidation, and supercapacitors. Materials 9: 1-15. |
| [20] | Yu J, Wang D, Huang Y, et al. (2011) A cylindrical core-shell-like TiO2 nanotube array anode for flexible fiber-type dye-sensitized solar cells. Nanoscale Res Lett 6: 94. |
| [21] | Mondal K, Sharma A (2016) Recent advances in the synthesis and application of photocatalytic metal-metal oxide core-shell nanoparticles for environmental remediation and their recycling process. RSC Adv 6: 83589-83612. |
| [22] | Meng Y (2015) Synthesis and adsorption property of SiO2@Co(OH)2 core-shell nanoparticles. Nanomaterials 5: 554-564. |
| [23] | Jadhav J, Biswas S (2016) Structural and electrical properties of ZnO:Ag core shell nanoparticles synthesized by a polymer precursor method. Ceram Int 42: 16598-16610. |
| [24] | Addato SD, et al. (2015) Influence of size, shape and core-shell interface on surface plasmon resonance in Ag and Ag@MgO nanoparticle films deposited on Si/SiOx. Beilstein J Nanotechno 6: 404-413. |
| [25] | Muller A, Peglow S, Karnahl M, et al. (2018) Morphology, optical properties and photocatalytic activity of photo- and plasma-deposited Au and Au/Ag core/shell nanoparticles on titania layers. Nanomaterials 502: 6-12. |
| [26] | Senthilkumar N, Ganapathy M, Arulraj A, et al. (2018) Step synthesis of ZnO/Ag and ZnO/Au core/shell nanocomposites: Structural, optical and electrical property analysis. J Alloys Compd 750: 171-181. |
| [27] | Gawande MB, Goswami A, Asefa T, et al. (2015) Core-shell nanoparticles: Synthesis and applications in catalysis and electrocatalysis. Chem Soc Rev 44: 7540-7590. |
| [28] | Zhou M, Diao K, Zhang J, et al. (2014) Controllable synthesis of plasmonic ZnO/Au core/shell nanocable arrays on ITO glass. Physica E 56: 59-63. |
| [29] | Singh SC, Swarnkar RK, Gopal R (2010) Zn/ZnO core/shell nanoparticles synthesized by laser ablation in aqueous environment: Optical and structural characterizations. B Mater Sci 33: 21-26. |
| [30] | Oh S, Ha K, Kang S, et al. (2018) Self-standing ZnO nanotube/SiO2 core-shell arrays for high photon extraction efficiency in III-nitride emitter. Nanotechnology 28: 015301. |
| [31] | Beyene G, Senbeta T, Mesfin B (2019) Size dependent optical properties of ZnO@Ag core/shell nanostructures. Chinese J Phys 58: 235-243. |
| [32] | Brijitta J, Ramachandran D, Chennakesavulu K, et al. (2016) Mesoporous ZnO-SiO2 core-shell rods for uv absorbing and non-wetting applications. Mater Res Express 3: 25001. |
| [33] | Li F, Huang X, Jiang Y, Liu L, Li Z (2009) Synthesis and characterization of ZnO/SiO2 core/shell nanocomposites and hollow SiO2 nanostructures. Mater Res Bull 44: 437-441. |
| [34] | Agarwal S, Jangir LK, Rathore KS, et al. (2019) Morphology-dependent structural and optical properties of ZnO nanostructures. Appl Phys A-Mater 125: 553. |
| [35] | Getie S, Belay A, Chandra Reddy AR, et al. (2017) Synthesis and characterizations of zinc oxide nanoparticles for antibacterial applications. J Nanomed Nanotechno S 8: 004. |
| [36] | Pourshaban E, Abdizadeh H, Golobostanfard MR (2015) ZnO nanorods array synthesized by chemical bath deposition: effect of seed layer sol concentration. Procedia Mater Sci 11: 352-358. |
| [37] | Naeem-ur-Rehman M, Ali SM, Ramay SM, et al. (2019) Annealing induced defects in ZnO nanostructures. ApPhA 125: 528. |
| [38] | Hsu NF, Chang M, Hsu KT (2014) Rapid synthesis of ZnO dandelion-like nanostructures and their applications in humidity sensing and photocatalysis. Mater Sci Semicon Proc 21: 200-205. |
| [39] | Shi R, Yang P, Dong X, et al. (2013) Growth of flower-like ZnO on ZnO nanorod arrays created on zinc substrate through low-temperature hydrothermal synthesis. Appl Surf Sci 264: 162-170. |
| [40] | Koao LF, Dejene FB, Swart HC (2014) Properties of flower-like ZnO nanostructures synthesized using the chemical bath deposition. Mater Sci Semicon Proc 27: 33-40. |
| [41] | Ye J, Zhou R, Zheng C, et al. (2012) Size-controllable synthesis of spherical ZnO nanoparticles: Size- and concentration-dependent resonant light scattering. Microchem J 100: 61-65. |
| [42] | Chithra MJ, Pushpanathan K, Loganathan M. (2014) Structural and optical properties of Co-doped ZnO nanoparticles synthesized by precipitation method. Mater Manuf Process 29: 771-779. |
| [43] | Li P, Wei Y, Liu H, et al. (2005) Growth of well-defined ZnO microparticles with additives from aqueous solution. J Solid State Chem 178: 855-860. |
| [44] | Kołodziejczak-Radzimska A, Markiewicz E, Jesionowski T (2012) Structural characterization of ZnO particles obtained by the emulsion precipitation method. J Nanomater 2012: 1-9. |
| [45] | Ali MA, Idris MR, Quayum ME (2013) Fabrication of ZnO nanoparticles by solution combustion method for the photocatalytic degradation of organic dye. J Nanostructure Chem 3: 36. |
| [46] | Pu X, Zhang D, Yi X, et al. (2010) Rapid chemical synthesis and optical properties of ZnO ellipsoidal nanostructures. Adv Powder Technol 21: 344-349. |
| [47] | Wang Y, Wang B, Zhang Q, et al. (2013) Tunable electronic properties of ZnO nanowires and nanotubes under a transverse electric field. J Appl Phys 113: 034301. |
| [48] | Abdulkerim SYH, Yu HY, Wang C, et al. (2018) Sheet-like cellulose nanocrystal-ZnO nano-hybrids as multi-functional reinforcing agents in biopolyester composite nanofibers with ultrahigh UV-Shielding and antibacterial performances. ACS Appl Bio Mater 3: 714-727 |
| [49] | Ponnuvelu DV, Pullithadathil B, Prasad AK, et al. (2015) Rapid synthesis and characterization of hybrid ZnO@Au core shell nano rods for high performance, low temperature NO2 gas sensor applications. Appl Surf Sci 355: 726-735. |
| [50] | Azimi M, Sadjadi MS, Farhadyar N (2016) Fabrication and characterization of core/shell ZnO/gold nanostructures and study of their structural and optical properties. Orient J Chem 32: 2517-2523. |
| [51] | Noack J, Scheurell K, Kemnitz E, et al. (2012) MgF2 antireflective coatings by sol-gel processing: film preparation and thermal densification. J Mater Chem 22: 18535-18541. |
| [52] | Lö bmann P (2018) Sol-gel processing of MgF2 antireflective coatings. Nanomaterials 8: 295. |
| [53] | Ramesh Babu K, Lingam CB, Auluck S, et al. (2011) Structural, thermodynamic and optical properties of MgF2 studied from first principles theory. J Solid State Chem 184: 343-350. |
| [54] | Tan TT, Liu BJ, Wu ZH, et al. (2016) Annealing effects on structural, optical properties and laser-induced damage threshold of MgF2 thin films. Acta Metall Sin-Engl 30: 73-78. |
| [55] | Beyene G, Senbeta T, Mesfin B, et al. (2020) Plasmonic properties of spheroidal spindle and disc shaped core-shell nanostructures embedded in passive host matrices. Opt Quant Electron 52: 157. |
| [56] | Tanabe K (2016) A simple optical model well explains plasmonic-nanoparticle-enhanced spectral photocurrent in optically thin solar cells. Nanoscale Res Lett 11: 236. |
| [57] | Sambou A, Tall PD, Talla KH, at al. (2017) Control of the surface plasmon resonance of two configurations of nanoparticles: Simple gold nanorod and gold/Silica core/shell. Nanosci Nanotechnol Res 4: 1-6. |
| [58] | Piralaee M, Asgari A, Siahpoush V (2018) Plasmonic properties of spheroid silicon-silver nano-shells in prolate and oblate forms. Optik 172: 1064-1068. |
| [59] | Liu J, Su C, Ye Q, et al. (2018) Investigation of tunable surface plasmon resonances on spheroid core-shell alloy nanoparticles using DDA method. Preprints 2018070480. |
| [60] | Sambou A, Ngom BD, Gomis L, et al. (2016) Turnability of the plasmonic response of the gold nanoparticles in infrared region. Am J Nanomater 4: 63-69. |

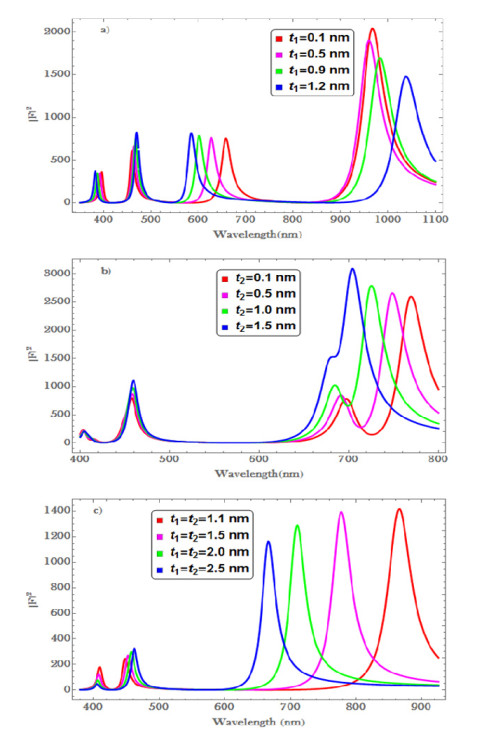
Figures(6) / Tables(1)

Gashaw Beyene, Gamachis Sakata, Teshome Senbeta, Belayneh Mesfin. Effect of core size/shape on the plasmonic response of spherical ZnO@Au core-shell nanostructures embedded in a passive host-matrices of MgF2[J]. AIMS Materials Science, 2020, 7(6): 705-719. doi: 10.3934/matersci.2020.6.705







 DownLoad:
DownLoad: