Citation: Siming Liu, Mengxin Wang, Yong Tan. Stabilizing inflation expectations in China: Does economic policy uncertainty matter?[J]. Green Finance, 2019, 1(4): 429-441. doi: 10.3934/GF.2019.4.429

| [1] |
Ang A, Bekaert G, Wei M (2007) Do macro variables, asset markets, or surveys forecast inflation better? J Monetary Econ 54: 1163-1212. Doi: 10.1016/j.jmoneco.2006.04.006 doi: 10.1016/j.jmoneco.2006.04.006

|
| [2] | Ankargren S, Unosson M, Yang Y (2018) A mixed-frequency Bayesian vector autoregression with a steady-state prior, Department of Statistics, Uppsala University. |
| [3] |
Baker SR, Bloom N, Davis SJ (2016) Measuring economic policy uncertainty. Q J Econ 131: 1593-1636. Doi:10.1093/qje/qjw024 doi: 10.1093/qje/qjw024
|
| [4] |
Caggiano G, Castelnuovo E, Groshenny N (2014) Uncertainty shocks and unemployment dynamics in U.S. recessions. J Monetary Econ 67: 78-92. Doi: 10.1016/j.jmoneco.2014.07.006 doi: 10.1016/j.jmoneco.2014.07.006
|
| [5] |
Caggiano G, Castelnuovo E, Figueres JM (2017) Economic policy uncertainty and unemployment in the United States: A nonlinear approach. Econ Lett 151: 31-34. Doi: 10.1016/j.econlet.2016.12.002 doi: 10.1016/j.econlet.2016.12.002
|
| [6] |
Carlson JA, Parkin M (1975) Inflation expectations. Economica 42: 123-138. Doi: 10.2307/2553588 doi: 10.2307/2553588
|
| [7] | Cerisola M, Gelos G (2009) What drives inflation expectations in Brazil?, an empirical analysis. Appl Econ 41: 1215-1227. Doi: 10.1080/00036840601166892 |
| [8] |
Chen L, Du Z, Tan Y (2019) Sustainable exchange rates in China: Is there the heterogeneous effect of economic policy uncertainty? Green Financ 1: 346-363.Doi:10.3934/GF.2019.4.346 doi: 10.3934/GF.2019.4.346
|
| [9] | Coibion O, Gorodnichenko Y, Kamdar R (2018a) The formation of expectations, inflation, and the Phillips Curve. J Econ Lit 56: 1447-1491. Doi: 10.1257/jel.20171300 |
| [10] | Coibion O, Gorodnichenko Y, Kumar S, et al. (2018b) Inflation expectations as a policy tool? NBER Working paper No. w24788. Doi: 10.3386/w24788 |
| [11] | Davis SJ (2016) An index of global economic policy uncertainty. NBER Working paper No. w22740. Doi: 10.3386/w22740 |
| [12] |
Ferrario A, Guidolin M, Pedio M (2018) Comparing in- and out-of-sample approaches to variance decomposition-based estimates of network connectedness an application to the Italian banking system. Quant Financ Econ 2: 661-701. Doi: 10.3934/QFE.2018.3.661 doi: 10.3934/QFE.2018.3.661
|
| [13] |
Fontaine I, Didier L, Razafindravaosolonirina J (2017) Foreign policy uncertainty shocks and US macroeconomic activity: Evidence from China. Econ Lett 155: 121-125. Doi: 10.1016/j.econlet.2017.03.034 doi: 10.1016/j.econlet.2017.03.034
|
| [14] |
Fukuda S, Teruyama H, Toda HY (1991) Inflation and price-wage dispersions in Japan. J Jpn Int Econ 5: 160-188. Doi: 10.1016/0889-1583(91)90020-Q doi: 10.1016/0889-1583(91)90020-Q
|
| [15] | Gauvin L, McLoughlin C, Reinhardt D (2013) Policy Uncertainty Spillovers to Emerging Markets-Evidence from Capital Flows. SSRN Electron J. Doi:10.2139/ssrn.2273452 |
| [16] | Ghosh T, Sahu S, Chattopadhyay S (2017) Households' inflation expectations in India: Role of economic policy uncertainty and global financial uncertainty spill-over. Indira Gandhi Institute of Development Research, Mumbai, India. |
| [17] |
Ghysels E (2016) Macroeconomics and the reality of mixed frequency data. J Econometrics 193: 294-314. Doi: 10.1016/j.jeconom.2016.04.008 doi: 10.1016/j.jeconom.2016.04.008
|
| [18] |
Globan T, Arčabić V, Sorić P (2016) Inflation in new EU member states: a domestically or externally driven phenomenon? Emerg Mark Financ Trade 52: 154-168. Doi: 10.1080/1540496X.2014.998547 doi: 10.1080/1540496X.2014.998547
|
| [19] | Gürkaynak RS, Swanson E, Levin A (2010) Does inflation targeting anchor long-run inflation expectations? evidence from the U.S., UK, and Sweden. J Eur Econ Assoc 8: 1208-1242. Doi: 10.1162/jeea_a_00023 |
| [20] |
Hachula M, Nautz D (2018) The dynamic impact of macroeconomic news on long-term inflation expectations. Econ Lett 165: 39-43. Doi: 10.1016/j.econlet.2018.01.015 doi: 10.1016/j.econlet.2018.01.015
|
| [21] |
Hammoudeh S, Reboredo JC (2018) Oil price dynamics and market-based inflation expectations. Energy Econ 75: 484-491. Doi: 10.1016/j.eneco.2018.09.011 doi: 10.1016/j.eneco.2018.09.011
|
| [22] |
Han L, Qi M, Yin L (2016) Macroeconomic policy uncertainty shocks on the Chinese economy: a GVAR analysis. Appl Econ 48: 4907-4921. Doi:10.1080/00036846.2016.1167828 doi: 10.1080/00036846.2016.1167828
|
| [23] | Istrefi K, Piloiu P (2014) Economic policy uncertainty and inflation expectations. Banque de France Working Paper. No. 511. Doi: 10.2139/ssrn.2510829 |
| [24] |
Kang WS, de Gracia FP, Ratti RA (2017) Oil price shocks, policy uncertainty and stock returns of oil and gas corporations. J Int Money Financ 70: 344-359. Doi: 10.1016/j.jimonfin.2016.10.003 doi: 10.1016/j.jimonfin.2016.10.003
|
| [25] |
Lee YM, Wang KM (2017) How do economic growth asymmetry and inflation expectations affect Fisher hypothesis and Fama's proxy hypothesis? Quant Financ Econ 1: 428-453. Doi: 10.3934/QFE.2017.4.428 doi: 10.3934/QFE.2017.4.428
|
| [26] |
Lei CY, Lu Z, Zhang CS (2015) News on inflation and the epidemiology of inflation expectations in China? Econ Syst 39: 644-453. Doi: 10.1016/j.ecosys.2015.04.006 doi: 10.1016/j.ecosys.2015.04.006
|
| [27] | Li Z, Zhong J (2019) Impact of economic policy uncertainty shocks on China's financial conditions. Financ Res Lett, [in press]. Doi: 10.1016/j.frl.2019.101303 |
| [28] |
Malmendier U, Nagel S (2016) Learning from inflation experiences. Q J Econ 131: 53-87. Doi:10.1093/qje/qjw037 doi: 10.1093/qje/qjv037
|
| [29] | Mullineaux DJ (1980) Inflation expectations and money growth in the United States. Am Econ Rev 70: 149-161. URL:https://www.jstor.org/stable/1814744 |
| [30] |
Pearce DK (1987) Short-term inflation expectations: Evidence from a monthly survey: note. J Money Credit Bank 19: 388-395. Doi: 10.2307/1992084 doi: 10.2307/1992084
|
| [31] |
Pesaran MH (1985) Formation of inflation expectations in British manufacturing industries. Econ J 95: 948-975. Doi: 10.2307/2233258 doi: 10.2307/2233258
|
| [32] |
Schorfheide F, Song D (2015) Real-time forecasting with a mixed-frequency VAR. J Bus Econ Stat 33: 366-380. Doi: 10.1080/07350015.2014.954707 doi: 10.1080/07350015.2014.954707
|
| [33] |
Serdar O, Ismet G (2019) Re-considering the Fisher equation for South Korea in the application of nonlinear and linear ARDL models. Quant Financ Econ 3: 75-87. Doi: 10.3934/QFE.2019.1.75 doi: 10.3934/QFE.2019.1.75
|
| [34] |
Stockhammar P, Österholm P (2015) Effects of US policy uncertainty on Swedish GDP growth. Empir Econ 50: 443-462. Doi:10.1007/s00181-015-0934-y doi: 10.1007/s00181-015-0934-y
|
| [35] |
Szyszko M, Płuciennik P (2018) Factors driving consumers' inflation expectations: does the central bank's inflation forecast really matter? Eastern Eur Econ 56: 307-328. Doi: 10.1080/00128775.2018.1467781 doi: 10.1080/00128775.2018.1467781
|
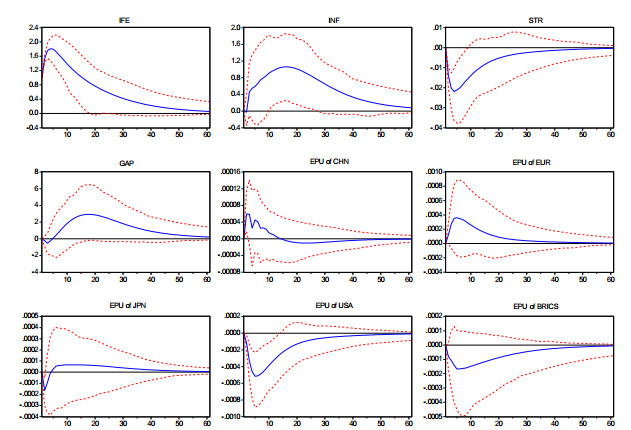
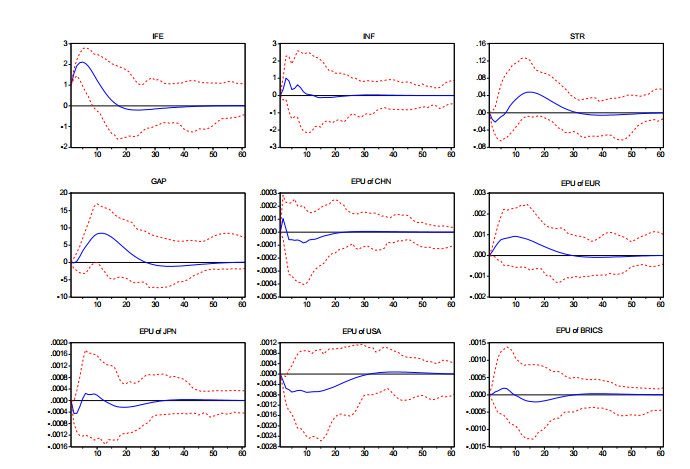
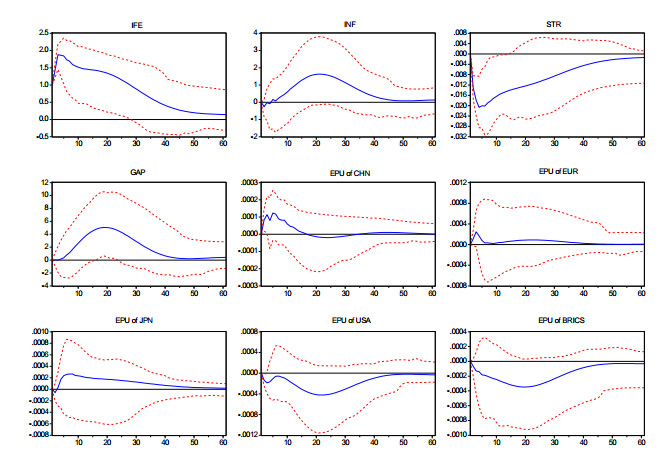
Figures(3) / Tables(4)

Siming Liu, Mengxin Wang, Yong Tan. Stabilizing inflation expectations in China: Does economic policy uncertainty matter?[J]. Green Finance, 2019, 1(4): 429-441. doi: 10.3934/GF.2019.4.429







 DownLoad:
DownLoad: