Citation: Simone Göttlich, Stephan Knapp. Modeling random traffic accidents by conservation laws[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1677-1701. doi: 10.3934/mbe.2020088

| [1] | M. Garavello, K. Han, B. Piccoli, Models for vehicular traffic on networks, vol. 9 of AIMS Series on Applied Mathematics, American Institute of Mathematical Sciences (AIMS), Springfield, MO, 2016. |
| [2] | M. Garavello, B. Piccoli, Traffic flow on networks, vol. 1 of AIMS Series on Applied Mathematics, American Institute of Mathematical Sciences (AIMS), Springfield, MO, 2006, Conservation laws models. |
| [3] | S. Blandin, P. Goatin, Well-posedness of a conservation law with non-local flux arising in traffic flow modeling, Numer. Math., 132 (2016), 217-241. |
| [4] | R. M. Colombo, Hyperbolic phase transitions in traffic flow, SIAM J. Appl. Math., 63 (2002), 708-721. |
| [5] | H. Holden, N. H. Risebro, A mathematical model of traffic flow on a network of unidirectional roads, SIAM J. Math. Anal., 26 (1995), 999-1017. |
| [6] | M. Gugat, M. Herty, A. Klar, G. Leugering, Optimal control for traffic flow networks, J. Optim. Theory Appl., 126 (2005), 589-616. |
| [7] | S. Fan, M. Herty, B. Seibold, Comparative model accuracy of a data-fitted generalized Aw-RascleZhang model, Netw. Heterog. Media, 9 (2014), 239-268. |
| [8] | S. E. Jabari, H. X. Liu, A stochastic model of traffic flow: Gaussian approximation and estimation, Transport. Res. B-Meth., 47 (2013), 15-41. |
| [9] | A. Sopasakis, M. A. Katsoulakis, Stochastic modeling and simulation of traffic flow: asymmetric single exclusion process with Arrhenius look-ahead dynamics, SIAM J. Appl. Math., 66 (2006), 921-944. |
| [10] | M. Herty, V. Schleper, Traffic flow with unobservant drivers, ZAMM Z. Angew. Math. Mech., 91 (2011), 763-776. |
| [11] | S. Moutari, M. Herty, A Lagrangian approach for modeling road collisions using second-order models of traffic flow, Commun. Math. Sci., 12 (2014), 1239-1256. |
| [12] | S. Moutari, M. Herty, A. Klein, M. Oeser, B. Steinauer, V. Schleper, Modelling road traffic accidents using macroscopic second-order models of traffic flow, IMA J. Appl. Math., 78 (2013), 1087-1108. |
| [13] | L. Jin, S. Amin, Analysis of a stochastic switching model of freeway traffic incidents, IEEE Trans. Automat. Control, 64 (2019), 1093-1108. |
| [14] | M. Baykal-Gürsoy, W. Xiao, K. Ozbay, Modeling traffic flow interrupted by incidents, Eur. J. Oper. Res., 195 (2009), 127-138. |
| [15] | P. Freguglia, A. Tosin, Proposal of a risk model for vehicular traffic: a Boltzmann-type kinetic approach, Commun. Math. Sci., 15 (2017), 213-236. |
| [16] | M. J. Lighthill, G. B. Whitham, On kinematic waves. II. A theory of traffic flow on long crowded roads, Proc. Roy. Soc. London. Ser. A., 229 (1955), 317-345. |
| [17] | M. H. A. Davis, Piecewise-deterministic Markov processes: a general class of nondiffusion stochastic models, J. Roy. Statist. Soc. Ser. B, 46 (1984), 353-388. |
| [18] | M. Jacobsen, Point Process Theory and Applications, Probability and its Applications, Birkhäuser Boston, Inc., Boston, MA, 2006, Marked point and piecewise deterministic processes. |
| [19] | S. Göttlich, S. Knapp, Load-Dependent Machine Failures in Production Network Models, SIAM J. Appl. Math., 79 (2019), 1197-1217. |
| [20] | A. Barth, F. G. Fuchs, Uncertainty quantification for hyperbolic conservation laws with flux coefficients given by spatiotemporal random fields, SIAM J. Sci. Comput., 38 (2016), A2209-A2231. |
| [21] | S. Mishra, N. H. Risebro, C. Schwab, S. Tokareva, Numerical solution of scalar conservation laws with random flux functions, SIAM/ASA J. Uncertain. Quan., 4 (2016), 552-591. |
| [22] | U. S. Fjordholm, S. Lanthaler, S. Mishra, Statistical solutions of hyperbolic conservation laws: foundations, Arch. Ration. Mech. An., 226 (2017), 809-849. |
| [23] | G. M. Coclite, N. H. Risebro, Conservation laws with time dependent discontinuous coefficients, SIAM J. Math. Anal., 36 (2005), 1293-1309. |
| [24] | H. Holden, N. H. Risebro, Front Tracking for Hyperbolic Conservation Laws, vol. 152 of AMS, 2nd edition, Springer, Heidelberg, 2015. |
| [25] | K. H. Karlsen, J. D. Towers, Convergence of the Lax-Friedrichs scheme and stability for conservation laws with a discontinuous space-time dependent flux, Chinese Ann. Math., 25 (2004), 287-318. |
| [26] | J. D. Towers, Convergence of a difference scheme for conservation laws with a discontinuous flux, SIAM J. Numer. Anal., 38 (2000), 681-698. |
| [27] | J. D. Towers, Convergence via OSLC of the Godunov scheme for a scalar conservation law with time and space flux discontinuities, Numer. Math., 139 (2018), 939-969. |
| [28] | H. Bauer, Measure and Integration Theory, vol. 26 of De Gruyter Studies in Mathematics, Walter de Gruyter & Co., Berlin, 2001. Available from: https://doi.org/10.1515/9783110866209, Translated from the German by Robert B. Burckel. |
| [29] | L. C. Evans, R. F. Gariepy, Measure theory and fine properties of functions, Revised edition, Textbooks in Mathematics, CRC Press, Boca Raton, FL, 2015. |
| [30] | P. R. Halmos, Measure Theory, Springer New York, 1978. Available from: https://www.ebook.de/de/product/3686564/paul_r_halmos_measure_theory.html. |
| [31] | W. Rudin, Real and Complex Analysis, 3rd edition, McGraw-Hill Book Co., New York, 1987. |
| [32] | V. Lemaire, M. Thieullen, N. Thomas, Exact simulation of the jump times of a class of piecewise deterministic Markov processes, J. Sci. Comput., 75 (2018), 1776-1807. |
| [33] | P. Degond, C. Ringhofer, Stochastic dynamics of long supply chains with random breakdowns, SIAM J. Appl. Math., 68 (2007), 59-79. |
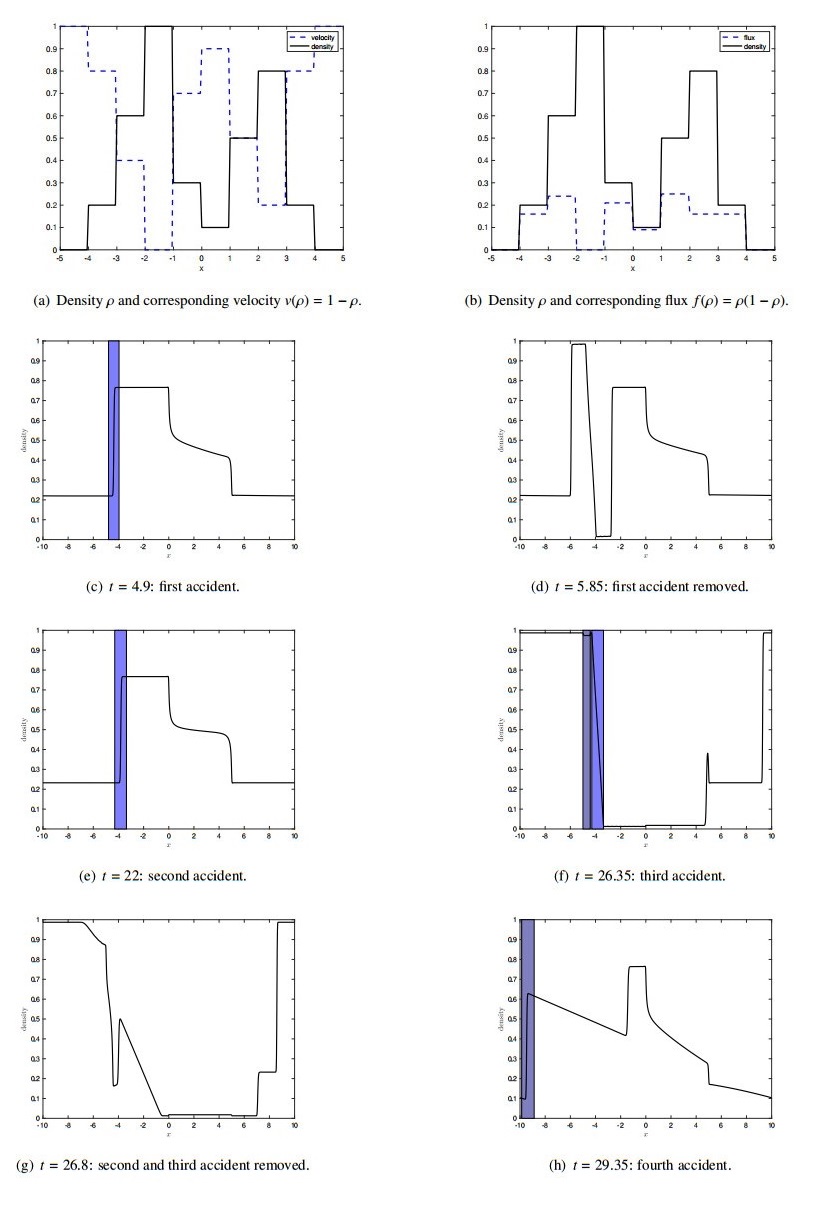
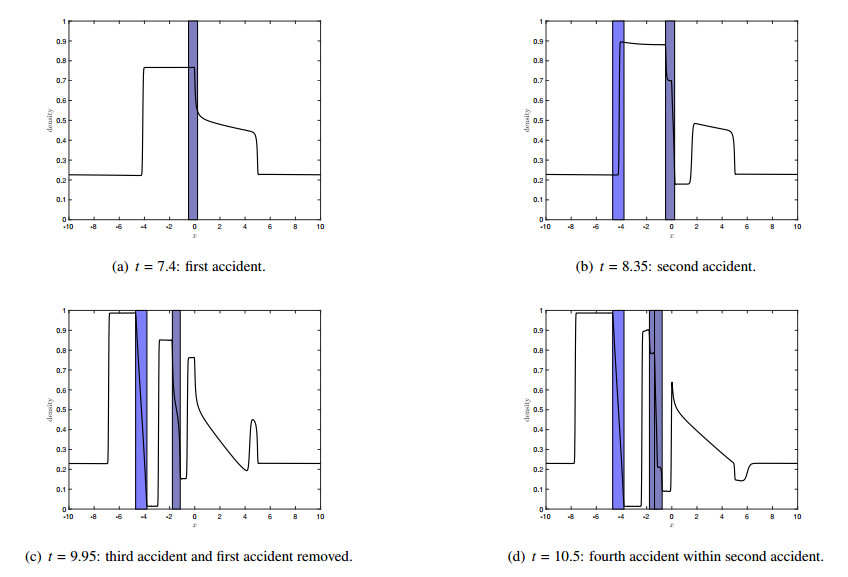
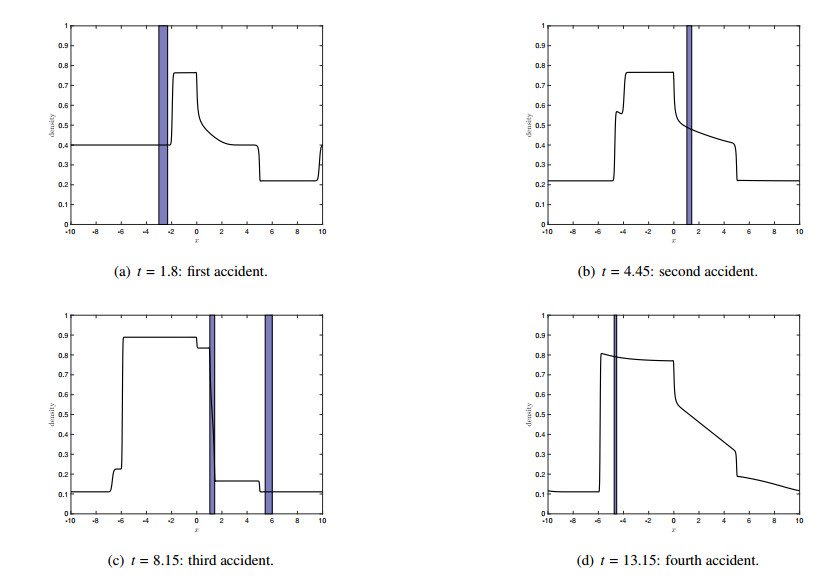
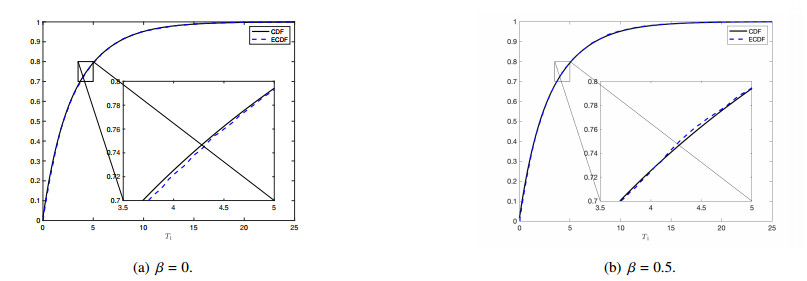
Figures(7)

Simone Göttlich, Stephan Knapp. Modeling random traffic accidents by conservation laws[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1677-1701. doi: 10.3934/mbe.2020088







 DownLoad:
DownLoad: