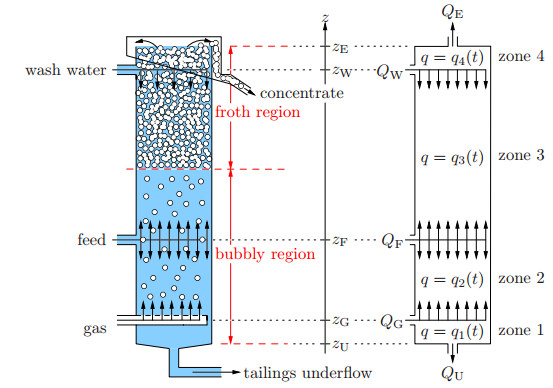
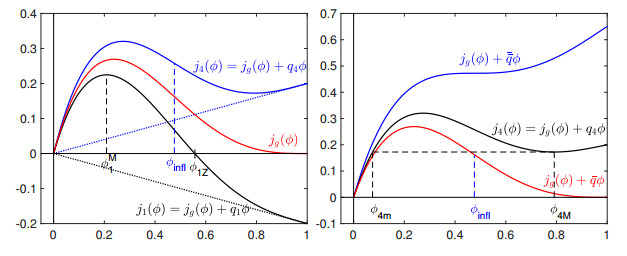
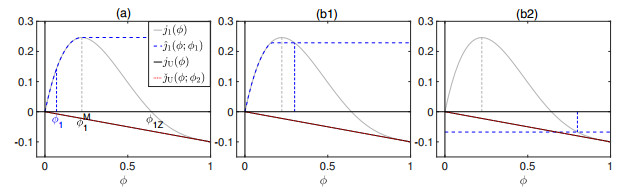
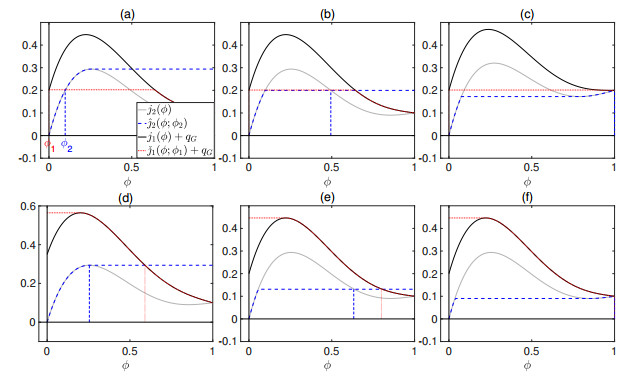
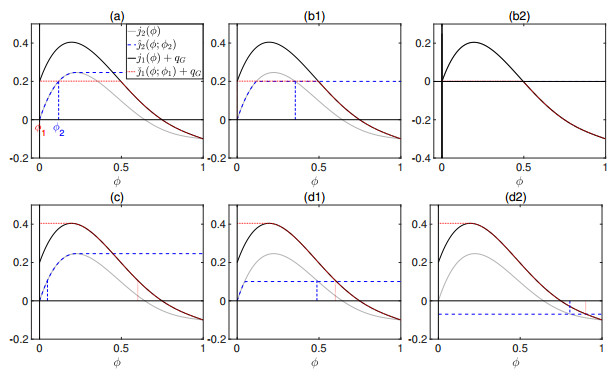
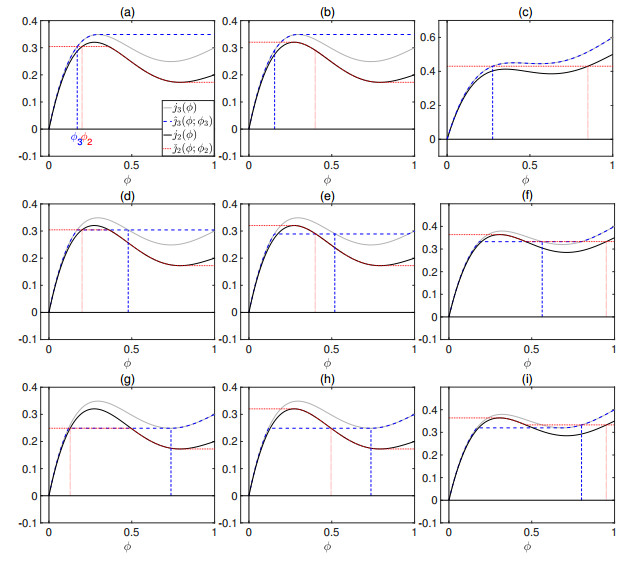
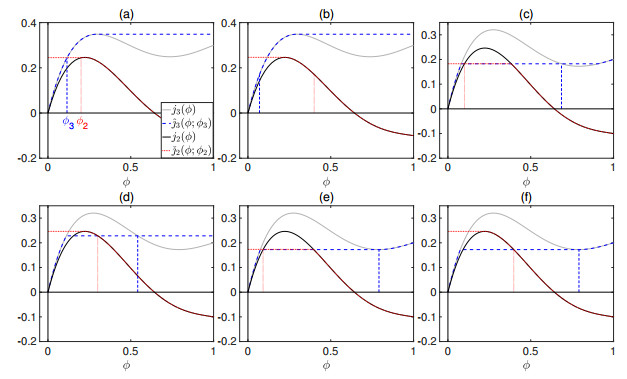
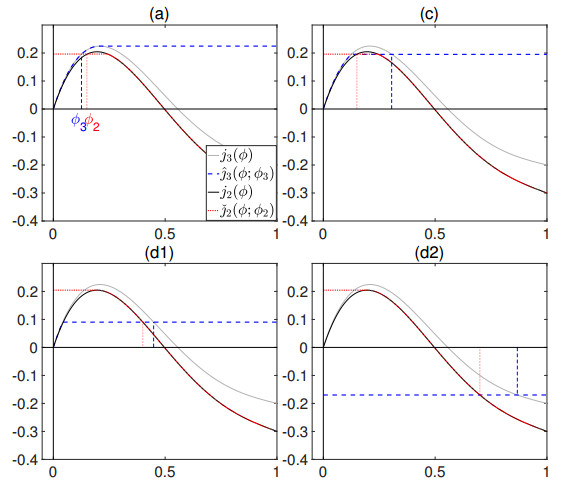
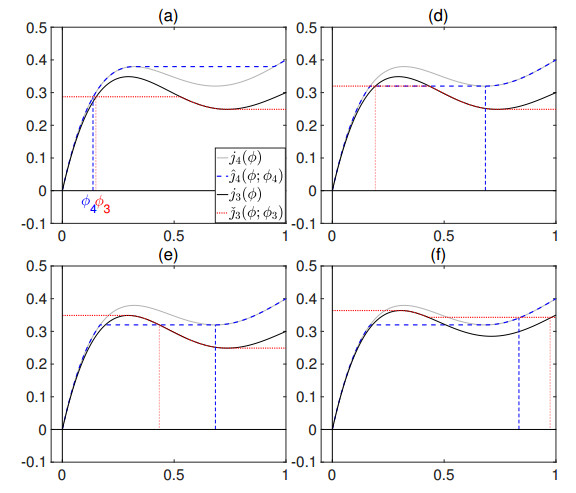
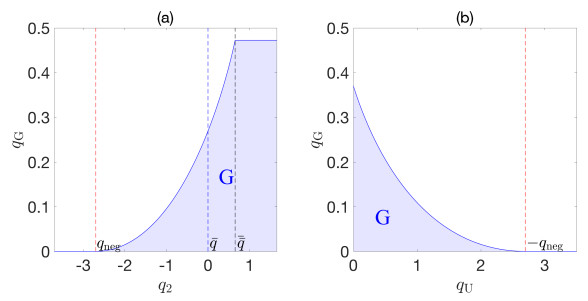
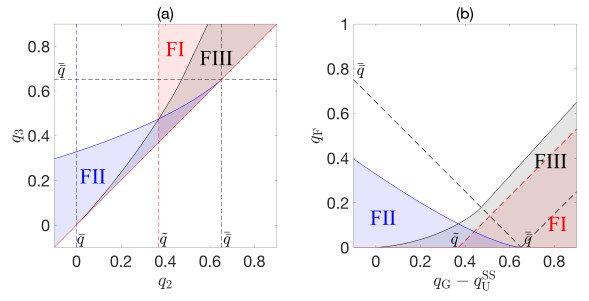
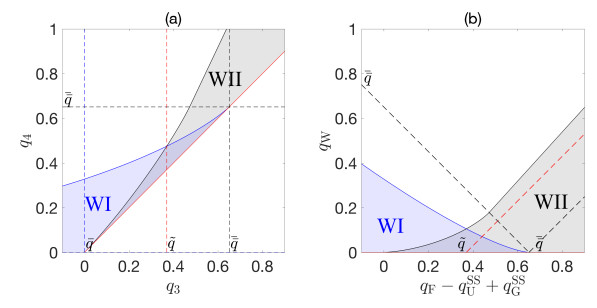
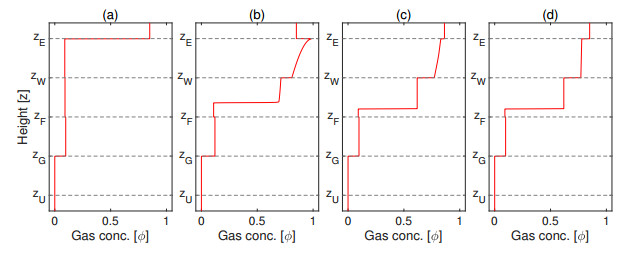
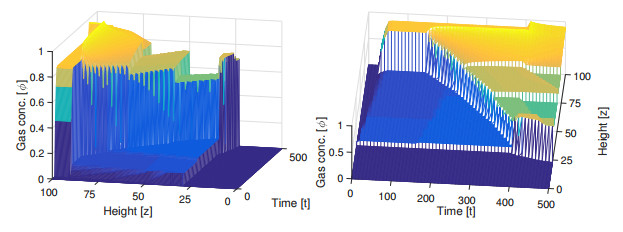
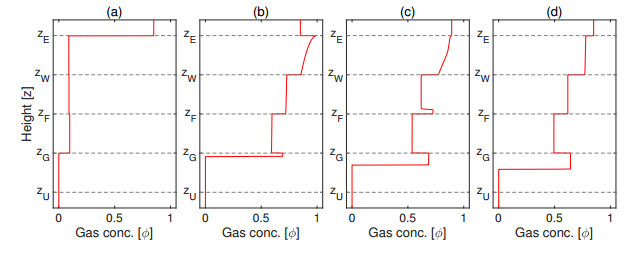
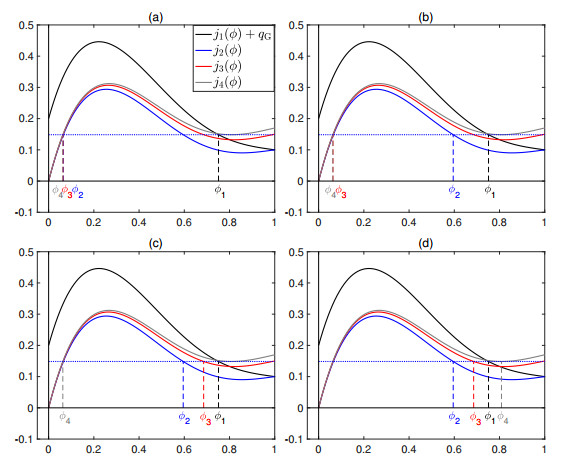
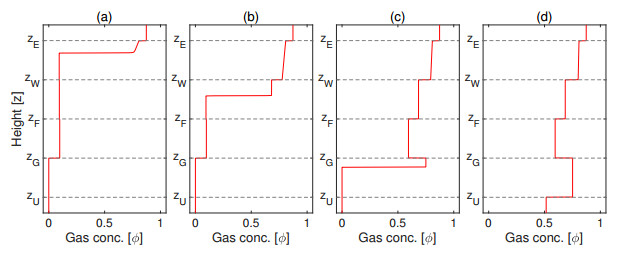
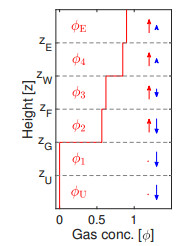
Flotation is a unit operation extensively used in the recovery of valuable minerals in mineral processing and related applications. Essential insight to the hydrodynamics of a flotation column can be obtained by studying just two phases: gas and fluid. To this end, the approach based on the drift-flux theory, proposed in similar form by several authors, is reformulated as a one-dimensional non-linear conservation law with a multiply discontinuous flux. The unknown is the gas volume fraction as a function of height and time, and the flux function depends discontinuously on spatial position due to several feed inlets. The resulting model is similar, but not equivalent, to previously studied clarifier-thickener models for solid-liquid separation and therefore adds a new real-world application to the field of conservation laws with discontinuous flux. Steady-state solutions are studied in detail, including their construction by applying an appropriate entropy condition across each flux discontinuity. This analysis leads to operating charts and tables collecting all possible steady states along with some necessary conditions for their feasibility in each case. Numerical experiments show that the transient model recovers the steady states, depending on the feed rates of the different inlets.
Citation: Raimund Bürger, Stefan Diehl, María Carmen Martí. A conservation law with multiply discontinuous flux modelling a flotation column[J]. Networks and Heterogeneous Media, 2018, 13(2): 339-371. doi: 10.3934/nhm.2018015
Flotation is a unit operation extensively used in the recovery of valuable minerals in mineral processing and related applications. Essential insight to the hydrodynamics of a flotation column can be obtained by studying just two phases: gas and fluid. To this end, the approach based on the drift-flux theory, proposed in similar form by several authors, is reformulated as a one-dimensional non-linear conservation law with a multiply discontinuous flux. The unknown is the gas volume fraction as a function of height and time, and the flux function depends discontinuously on spatial position due to several feed inlets. The resulting model is similar, but not equivalent, to previously studied clarifier-thickener models for solid-liquid separation and therefore adds a new real-world application to the field of conservation laws with discontinuous flux. Steady-state solutions are studied in detail, including their construction by applying an appropriate entropy condition across each flux discontinuity. This analysis leads to operating charts and tables collecting all possible steady states along with some necessary conditions for their feasibility in each case. Numerical experiments show that the transient model recovers the steady states, depending on the feed rates of the different inlets.

| [1] |
A theory of 
|
| [2] | O. A. Bascur, Example of a dynamic flotation framework, In: Centenary of Flotation Symposium, Brisbane, QLD, 6-9 June 2005, Australasian Institute of Mining and Metallurgy Publication Series, 2005, 85-91. |
| [3] | A consistent modelling methodology for secondary settling tanks: A reliable numerical method. Water Sci. Technol. (2013) 68: 192-208. |
| [4] | R. Bürger, S. Diehl and C. Mejías, A difference scheme for a degenerating convection-diffusion-reaction system modelling continuous sedimentation, ESAIM: Math. Model. Numer. Anal., to appear. |
| [5] |
Conservation laws with discontinuous flux: A short introduction. J. Eng. Math. (2008) 60: 241-247.
|
| [6] |
On an extended clarifier-thickener model with singular source and sink terms. Eur. J. Appl. Math. (2006) 17: 257-292.
|
| [7] |
A family of numerical schemes for kinematic flows with discontinuous flux. J. Eng. Math. (2008) 60: 387-425.
|
| [8] |
Well-posedness in
|
| [9] |
Second-order schemes for conservation laws with discontinuous flux modelling clarifier-thickener units. Numer. Math. (2010) 116: 579-617.
|
| [10] |
A model of continuous sedimentation of flocculated suspensions in clarifier-thickener units. SIAM J. Appl. Math. (2005) 65: 882-940.
|
| [11] |
On some difference schemes and entropy conditions for a class of multi-species kinematic flow models with discontinuous flux. Netw. Heterog. Media (2010) 5: 461-485.
|
| [12] | J. M. Coulson, J. F. Richardson, J. R. Backhurst and J. H. Harker, Coulson and Richardson's Chemical Engineering. Volume 2: Particle Technology and Separation Processes, Fourth Ed., Butterworth-Heinemann, Oxford, 2000. |
| [13] |
Fluidized bed desliming in fine particle flotation, Part Ⅰ. Chem. Eng. Sci. (2014) 108: 283-298.
|
| [14] |
On scalar conservation laws with point source and discontinuous flux function. SIAM J. Math. Anal. (1995) 26: 1425-1451.
|
| [15] |
A conservation law with point source and discontinuous flux function modelling continuous sedimentation. SIAM J. Appl. Math. (1996) 56: 388-419.
|
| [16] |
Operating charts for continuous sedimentation Ⅰ: Control of steady states. J. Eng. Math. (2001) 41: 117-144.
|
| [17] |
Operating charts for continuous sedimentation Ⅱ: Step responses. J. Eng. Math. (2005) 53: 139-185.
|
| [18] |
A uniqueness condition for nonlinear convection-diffusion equations with discontinuous coefficients. J. Hyperbolic Differential Equations (2009) 6: 127-159.
|
| [19] |
Numerical identification of constitutive functions in scalar nonlinear convection-diffusion equations with application to batch sedimentation. Appl. Numer. Math. (2015) 95: 154-172.
|
| [20] |
Fast reliable simulations of secondary settling tanks in wastewater treatment with semi-implicit time discretization. Comput. Math. Appl. (2015) 70: 459-477.
|
| [21] | J. A. Finch and G. S. Dobby, Column Flotation, Pergamon Press, London, 1990. |
| [22] |
bed desliming in fine particle flotation Part Ⅱ: Flotation of a model feed. Chem. Eng. Sci. (2014) 108: 299-309.
|
| [23] | Finite difference methods for numerical computation of discontinuous solutions of equations of fluid dynamics. Mat. Sb. (1959) 47: 271-306. |
| [24] |
H. Holden and N. H. Risebro, Front Tracking for Hyperbolic Conservation Laws, Second Edition, Springer Verlag, Berlin, 2015. doi: 10.1007/978-3-662-47507-2
|
| [25] | Liquid transport in multi-layer froths. J. Colloid Interf. Sci. (2007) 314: 207-213. |
| [26] | First order quasilinear equations in several independent variables. Math. USSR-Sb. (1970) 10: 217-243. |
| [27] |
The coexistence of the froth and liquid phases in a flotation column. Chem. Eng. Sci. (1992) 47: 4345-4355.
|
| [28] | S. Mishra, Numerical methods for conservation laws with discontinuous coefficients, Chapter 18 in R. Abgrall and C.-W. Shu (eds.), Handbook of Numerical Methods for Hyperbolic Problems: Applied and Modern Issues, North Holland, 18 (2017), 479-506. |
| [29] |
Analysis of creaming and formation of foam layer in aerated liquid. J. Colloid Interface Sci. (2010) 345: 566-572.
|
| [30] | O. A. Oleinik, Uniqueness and stability of the generalized solution of the Cauchy problem for a quasi-linear equation Uspekhi Mat. Nauk., 14 (1959), 165-170. Amer. Math. Soc. Trans. Ser. 2, 33 (1964), 285-290. |
| [31] |
Flow characterization of a flotation column. Canad. J. Chem. Eng. (1989) 67: 916-923.
|
| [32] |
Oil recovery from oil in water emulsions using a flotation column. Canad. J. Chem. Eng. (1990) 68: 959-967.
|
| [33] | Amenability testing of fine coal beneficiation using laboratory flotation column. Materials Transactions (2015) 56: 766-773. |
| [34] |
Sedimentation and fluidisation: Part Ⅰ. Chemical Engineering Research and Design (1997) 75: 82-100.
|
| [35] |
Science and technology of dispersed two-phase systems—Ⅰ and Ⅱ. Chem. Eng. Sci. (1982) 37: 1125-1150.
|
| [36] |
A. Rushton, A. S. Ward and R. G. Holdich, Solid-Liquid Filtration and Separation Technology, Wiley Online Library, 2008. doi: 10.1002/9783527614974
|
| [37] | Convective-dispersive gangue transport in flotation froth. Chem. Eng. Sci. (2007) 62: 5736-5744. |
| [38] |
On the drift-flux analysis of flotation and foam fractionation processes. Canad. J. Chem. Eng. (2008) 86: 635-642.
|
| [39] | Drift flux modelling for a two-phase system in a flotation column. Canad. J. Chem. Eng. (2005) 83: 169-176. |
| [40] | G. B. Wallis, One-Dimensional two-Phase Flow, McGraw-Hill, New York, 1969. |
| [41] | The terminal speed of single drops or bubbled in an infinite medium. Int. J. Multiphase Flow (1974) 1: 491-511. |
| [42] | B. A. Wills and T. J. Napier-Munn, Wills' Mineral Processing Technology, Seventh Edition, Butterworth-Heinemann, Oxford, 2006. |
| [43] |
Bubble size estimation in a bubble swarm. J. Colloid Interf. Sci. (1988) 126: 37-44.
|
| [44] | A gas-liquid drift-flux model for flotation columns. Minerals Eng. (1993) 6: 199-205. |
Figures(22) / Tables(4)
Raimund Bürger, Stefan Diehl, María Carmen Martí. A conservation law with multiply discontinuous flux modelling a flotation column[J]. Networks and Heterogeneous Media, 2018, 13(2): 339-371. doi: 10.3934/nhm.2018015







 DownLoad:
DownLoad: