Citation: Junjing Xiong, Xiong Li, Hao Wang. The survival analysis of a stochastic Lotka-Volterra competition model with a coexistence equilibrium[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2717-2737. doi: 10.3934/mbe.2019135

| [1] | J. D. Murray, Mathematical Biology, Springer-Verlag, Berlin, Heidelberg, 2002. |
| [2] | X. Mao, S. Sabais and E. Renshaw, Asymptotic behavior of stochastic Lotka-Volterra model, J. Math. Anal. Appl., 287 (2003), 141–156. |
| [3] | N. H. Du and V. H. Sam, Dynamics of a stochastic Lotka-Volterra model perturbed by white noise, J. Math. Anal. Appl., 324 (2006), 82–97. |
| [4] | D. Jiang, C. Ji, X. Li, et al., Analysis of autonomous Lotka-Volterra competition systems with random perturbation, J. Math. Anal. Appl., 390 (2012), 582–595. |
| [5] | M. Liu and K. Wang, Population dynamical behavior of Lotka-Volterra cooperative systems with random perturbations, Discrete Contin. Dyn. Syst., 33 (2013), 2495–2522. |
| [6] | R. Rudnicki, Long-time behaviour of a stochastic prey-predator model, Stochastic Process. Appl., 108 (2003), 93–107. |
| [7] | D. H. Nguyen, N. H. Du and T. V. Ton, Asymptotic behavior of predator-prey systems perturbed by white noise, Acta Appl. Math., 115 (2011), 351–370. |
| [8] | D. H. Nguyen and G. Yin, Coexistence and exclusion of stochastic competitive Lotka-Volterra models, J. Differ. Equations, 262 (2017), 1192–1225. |
| [9] | M. Liu, K. Wang and Q. Wu, Survival analysis of stochastic competitive models in a polluted environment and stochastic competitive exclusion principle, Bull. Math. Biol., 73 (2011), 1969– 2012. |
| [10] | X. Mao, Stationary distribution of stochastic population systems, Syst. Control Lett., 60 (2011), 398–405. |
| [11] | A. Bahar and X. Mao, Stochastic delay Lotka-Volterra model, J. Math. Anal. Appl., 292 (2004), 364–380. |
| [12] | Q. Liu, The effects of time-dependent delays on global stability of stochastic Lotka-Volterra competitive model, Physica A., 420 (2015), 108–115. |
| [13] | J. J. Xiong, X. Li and H.Wang, Global asymptotic stability of a Lotka-Volterra competition model with stochasticity in inter-specific competition, Appl. Math. Letters., 89 (2019), 58–63. |
| [14] | A. V. Skorokhod, Asymptotic Methods in the Theory of Stochastic Differential Equations, American Mathematical Society, Providence, 1989. |
| [15] | L. R. Bellet, Ergodic properties of Markov processes, in Open Quantum System II, Springer, Berlin, Heidelberg, (2006), 1–39. |
| [16] | D. J. Higham, An algorithmic introduction to numerical simulation of stochastic differential equations, SIAM Rev., 43 (2001), 525–546. |
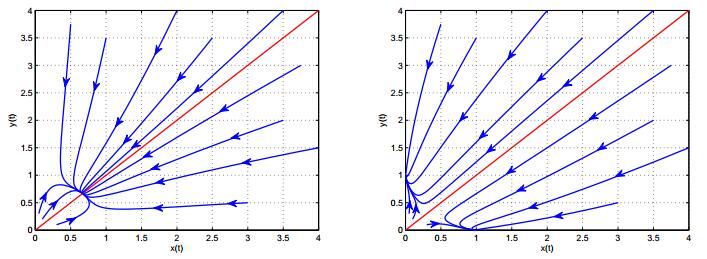
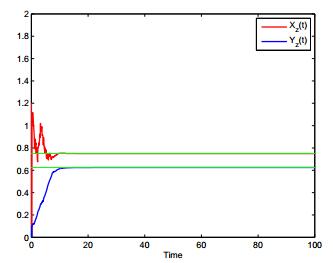
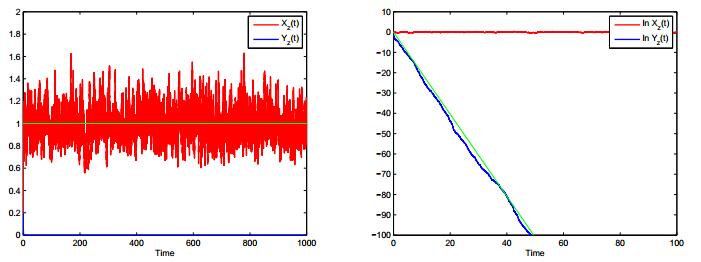
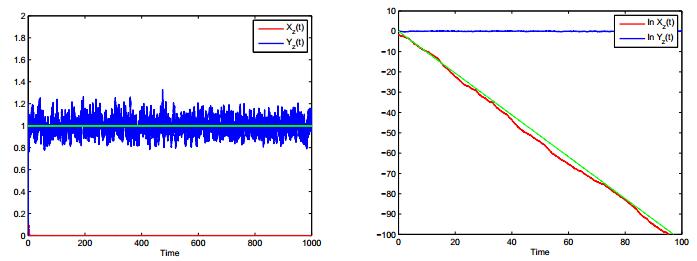
Figures(4)

Junjing Xiong, Xiong Li, Hao Wang. The survival analysis of a stochastic Lotka-Volterra competition model with a coexistence equilibrium[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2717-2737. doi: 10.3934/mbe.2019135







 DownLoad:
DownLoad: