Citation: Lucía Guadamuro, M. Andrea Azcárate-Peril, Rafael Tojo, Baltasar Mayo, Susana Delgado. Use of high throughput amplicon sequencing and ethidium monoazide dye to track microbiota changes in an equol-producing menopausal woman receiving a long-term isoflavones treatment[J]. AIMS Microbiology, 2019, 5(1): 102-116. doi: 10.3934/microbiol.2019.1.102

| [1] |
Xiao CW (2008) Health effects of soy protein and isoflavones in humans. J Nutr 138: 1244S–1249S. doi: 10.1093/jn/138.6.1244S

|
| [2] | Bolanos R, Del Castillo A, Francia J (2010) Soy isoflavones versus placebo in the treatment of climacteric vasomotor symptoms: systematic review and meta-analysis. Menopause 17: 660–666. |
| [3] | North American Menopause Society (2011) The role of soy isoflavones in menopausal health: report of The North American Menopause Society/Wulf H. Utian Translational Science Symposium in Chicago, IL (October 2010). Menopause 18: 732–753. |
| [4] |
EFSA (European Food Science Authority) (2012) Scientific Opinion on the substantiation of health claims related to soy isoflavones and maintenance of bone mineral density (ID 1655) and reduction of vasomotor symptoms associated with menopause (ID 1654, 1704, 2140, 3093, 3154, 3590) (further assessment) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J 10:2847. doi: 10.2903/j.efsa.2012.2847
|
| [5] |
Crozier A, Jaganath IB, Clifford MN (2009) Dietary phenolics: chemistry, bioavailability and effects on health. Nat Prod Rep 26: 1001–1043. doi: 10.1039/b802662a
|
| [6] |
de Cremoux P, This P, Leclercq G, et al. (2010) Controversies concerning the use of phytoestrogens in menopause management: bioavailability and metabolism. Maturitas 65: 334–339. doi: 10.1016/j.maturitas.2009.12.019
|
| [7] |
Sánchez-Calvo JM, Rodríguez-Iglesias MA, Molinillo JMG, et al. (2013) Soy isoflavones and their relationship with microflora: beneficial effects on human health in equol producers. Phytochem Rev 12: 979–1000. doi: 10.1007/s11101-013-9329-x
|
| [8] |
Atkinson C, Frankenfeld CL, Lampe JW (2005) Gut bacterial metabolism of the soy isoflavone daidzein: exploring the relevance to human health. Exp Biol Med (Maywood) 230: 155–170. doi: 10.1177/153537020523000302
|
| [9] |
Messina M (2016) Soy and Health Update: Evaluation of the Clinical and Epidemiologic Literature. Nutrients 8: E754. doi: 10.3390/nu8120754
|
| [10] |
Setchell KD, Clerici C (2010) Equol: history, chemistry, and formation. J Nutr 140: 1355S–1362S. doi: 10.3945/jn.109.119776
|
| [11] |
Kemperman RA, Bolca S, Roger LC, et al. (2010) Novel approaches for analysing gut microbes and dietary polyphenols: challenges and opportunities. Microbiology 156: 3224–3231. doi: 10.1099/mic.0.042127-0
|
| [12] | Clavel T, Lepage P, Charrier C (2014) The Family Coriobacteriaceae. In: Rosenberg E, DeLong EF, Lory S, et al. The Prokaryotes, 11 Eds., Berlin: Springer, 201–238. |
| [13] | Dueñas M, Muñoz-González I, Cueva C, et al. (2015) A survey of modulation of gut microbiota by dietary polyphenols. Biomed Res Int 2015: 850902. |
| [14] |
Nakatsu CH, Armstrong A, Clavijo AP, et al. (2014) Fecal bacterial community changes associated with isoflavone metabolites in postmenopausal women after soy bar consumption. PLoS One 9: e108924. doi: 10.1371/journal.pone.0108924
|
| [15] |
Bolca S, Possemiers S, Herregat A, et al. (2007) Microbial and dietary factors are associated with the equol producer phenotype in healthy postmenopausal women. J Nutr 137: 2242–2246. doi: 10.1093/jn/137.10.2242
|
| [16] |
Clavel T, Fallani M, Lepage P, et al. (2005) Isoflavones and functional foods alter the dominant intestinal microbiota in postmenopausal women. J Nutr 135: 2786–2792. doi: 10.1093/jn/135.12.2786
|
| [17] | Guadamuro L, Delgado S, Redruello B, et al. (2015) Equol status and changes in fecal microbiota in menopausal women receiving long-term treatment for menopause symptoms with a soy-isoflavone concentrate. Front Microbiol 6: 777 |
| [18] | Nocker A, Cheung CY, Camper AK (2006) Comparison of propidium monoazide with ethidium monoazide for differentiation of live vs. dead bacteria by selective removal of DNA from dead cells. J Microbiol Methods 67: 310–320. |
| [19] |
Redruello B, Guadamuro L, Cuesta I, et al. (2015) A novel UHPLC method for the rapid and simultaneous determination of daidzein, genistein and equol in human urine. J Chromatogr B Analyt Technol Biomed Life Sci 1005: 1–8. doi: 10.1016/j.jchromb.2015.09.029
|
| [20] |
Zoetendal EG, Heilig HG, Klaassens ES, et al. (2006) Isolation of DNA from bacterial samples of the human gastrointestinal tract. Nat Protoc 1: 870–873 doi: 10.1038/nprot.2006.142
|
| [21] |
Young JP, Downer HL, Eardly BD (1991) Phylogeny of the phototrophic rhizobium strain BTAi1 by polymerase chain reaction-based sequencing of a 16S rRNA gene segment. J Bacteriol 173: 2271–2277. doi: 10.1128/jb.173.7.2271-2277.1991
|
| [22] |
Goecks J, Nekrutenko A, Taylor J, et al. (2010) Galaxy: a comprehensive approach for supporting accessible, reproducible, and transparent computational research in the life sciences. Genome Biol 11: R86. doi: 10.1186/gb-2010-11-8-r86
|
| [23] |
Edgar RC, Haas BJ, Clemente JC, et al. (2011) UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27: 2194–2200. doi: 10.1093/bioinformatics/btr381
|
| [24] |
Wang Q, Garrity GM, Tiedje JM, et al. (2007) Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl Environ Microbiol 73: 5261–5267. doi: 10.1128/AEM.00062-07
|
| [25] |
Moreno CE, Halffter G (2001) On the measure of sampling effort used in species accumulation curves. J Appl Ecol 38: 487–490. doi: 10.1046/j.1365-2664.2001.00590.x
|
| [26] |
Yarza P, Richter M, Peplies J, et al. (2008) The All-Species Living Tree project: a 16S rRNA-based phylogenetic tree of all sequenced type strains. Syst Appl Microbiol 31: 241–250. doi: 10.1016/j.syapm.2008.07.001
|
| [27] |
Li W, Godzik A (2006) Cd-hit: a fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 22: 1658–1659. doi: 10.1093/bioinformatics/btl158
|
| [28] | Graf D, Di Cagno R, Fak F, et al. (2015) Contribution of diet to the composition of the human gut microbiota. Microb Ecol Health Dis 26: 26164. |
| [29] |
David LA, Maurice CF, Carmody RN, et al. (2014) Diet rapidly and reproducibly alters the human gut microbiome. Nature 505: 559–563. doi: 10.1038/nature12820
|
| [30] |
Singh RK, Chang HW, Yan D, et al. (2017) Influence of diet on the gut microbiome and implications for human health. J Transl Med, 15: 73. doi: 10.1186/s12967-017-1175-y
|
| [31] |
van Duynhoven J, Vaughan EE, Jacobs DM, et al. (2011) Metabolic fate of polyphenols in the human superorganism. Proc Natl Acad Sci USA 108: 4531–4538. doi: 10.1073/pnas.1000098107
|
| [32] |
Rowland IR, Wiseman H, Sanders TA, et al. (2000) Interindividual variation in metabolism of soy isoflavones and lignans: influence of habitual diet on equol production by the gut microflora. Nutr Cancer 36: 27–32. doi: 10.1207/S15327914NC3601_5
|
| [33] |
Guadamuro L, Dohrmann AB, Tebbe CC, et al. (2017) Bacterial communities and metabolic activity of faecal cultures from equol producer and non-producer menopausal women under treatment with soy isoflavones. BMC Microbiol 17: 93. doi: 10.1186/s12866-017-1001-y
|
| [34] |
Meehan CJ, Beiko RG (2014) A phylogenomic view of ecological specialization in the Lachnospiraceae, a family of digestive tract-associated bacteria. Genome Biol Evol 6: 703–713. doi: 10.1093/gbe/evu050
|
| [35] | Rios-Covian D, Ruas-Madiedo P, Margolles A, et al. (2016) Intestinal short chain fatty acids and their link with diet and human health. Front Microbiol 7: 185. |
| [36] |
Kim MG, Lee HS (2009) Growth-inhibiting activities of phenethyl isothiocyanate and its derivatives against intestinal bacteria. J Food Sci 74: M467–471. doi: 10.1111/j.1750-3841.2009.01333.x
|
| [37] |
Sklenickova O, Flesar J, Kokoska L, et al. (2010) Selective growth inhibitory effect of biochanin A against intestinal tract colonizing bacteria. Molecules 15: 1270–1279. doi: 10.3390/molecules15031270
|
| [38] |
Vazquez L, Guadamuro L, Giganto F, et al. (2017) Development and use of a real-time quantitative PCR method for detecting and quantifying equol-producing bacteria in human faecal samples and slurry cultures. Front Microbiol 8: 1155. doi: 10.3389/fmicb.2017.01155
|
| [39] |
Wagner AO, Malin C, Knapp BA, et al. (2008) Removal of free extracellular DNA from environmental samples by ethidium monoazide and propidium monoazide. Appl Environ Microbiol 74: 2537–2539. doi: 10.1128/AEM.02288-07
|
| [40] | Nocker A, Richter-Heitmann T, Montijn R, et al. (2010) Discrimination between live and dead cellsin bacterial communities from environmental water samples analyzed by 454 pyrosequencing. Int Microbiol 13: 59–65. |
| [41] |
Hein I, Flekna G, Wagner M, et al. (2006) Possible errors in the interpretation of ethidium bromide and PicoGreen DNA staining results from ethidium monoazide-treated DNA. Appl Environ Microbiol 72: 6860–6861. doi: 10.1128/AEM.01243-06
|
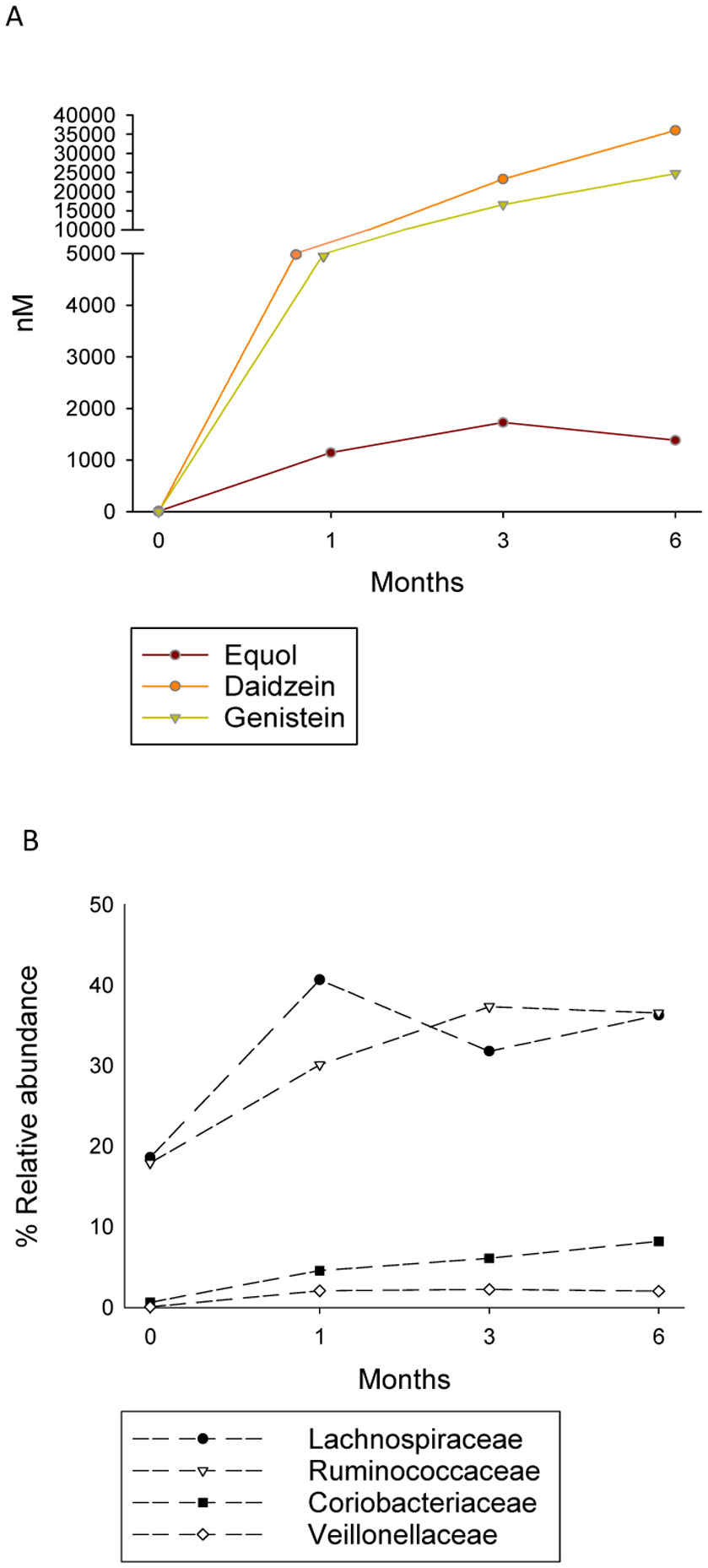
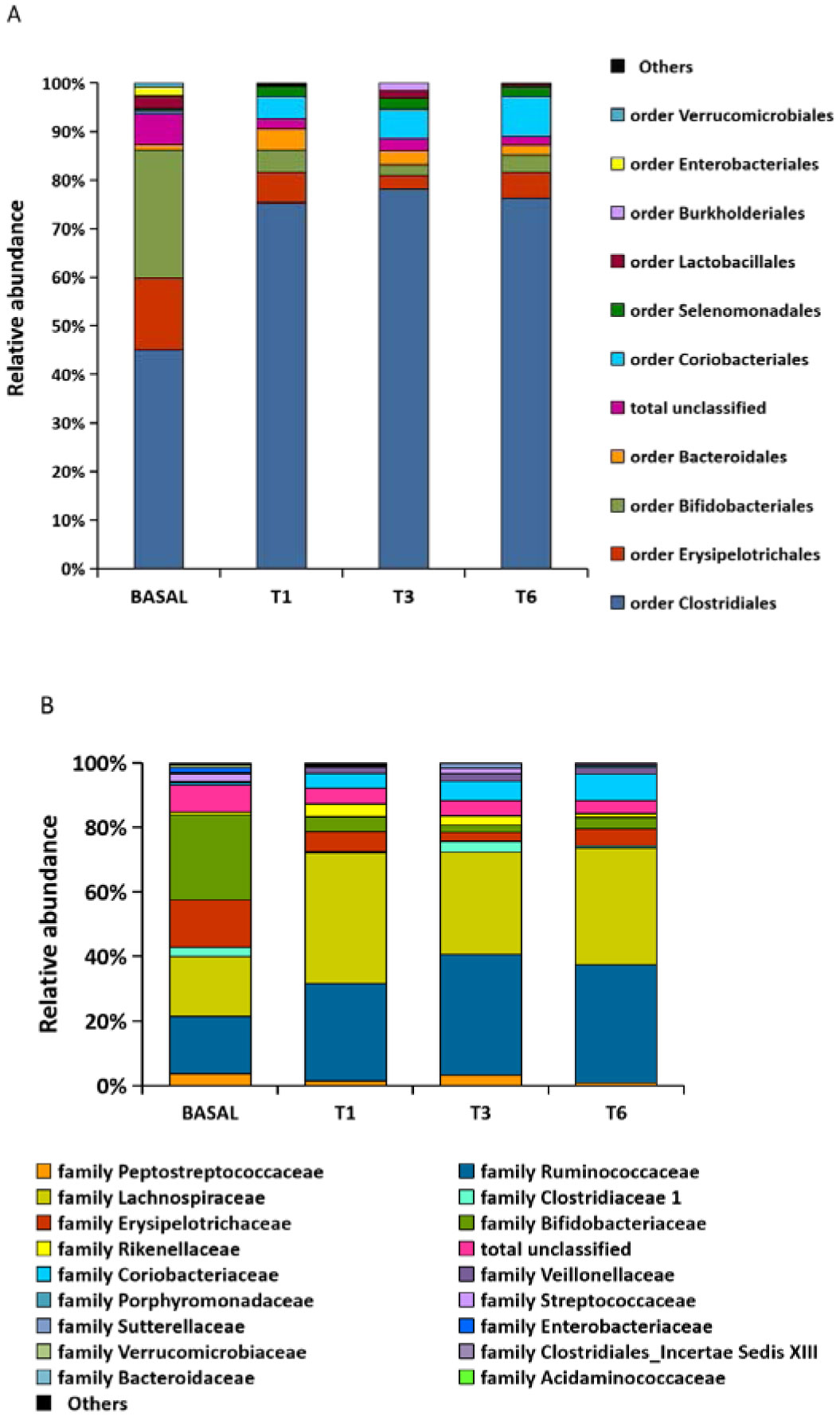
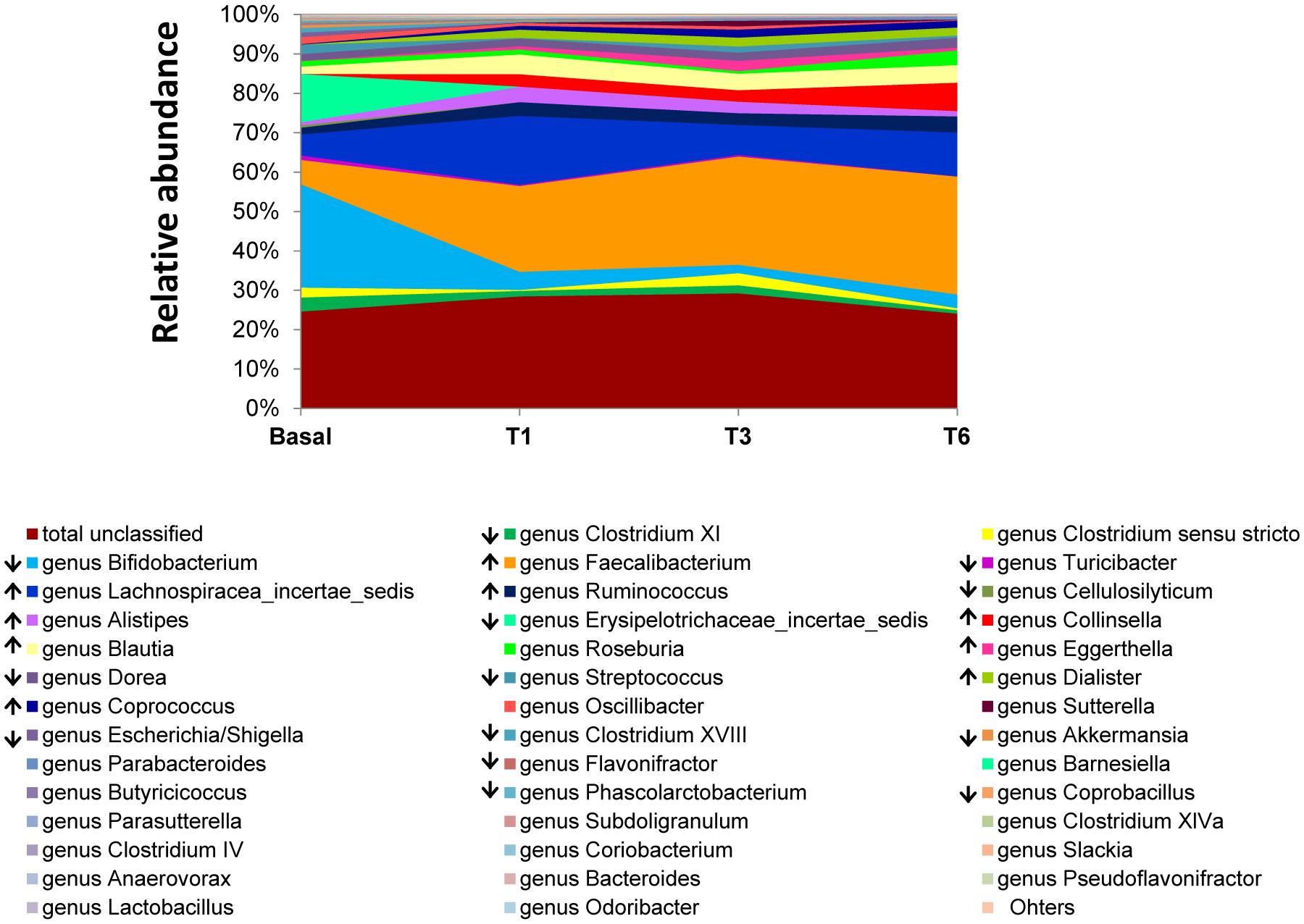
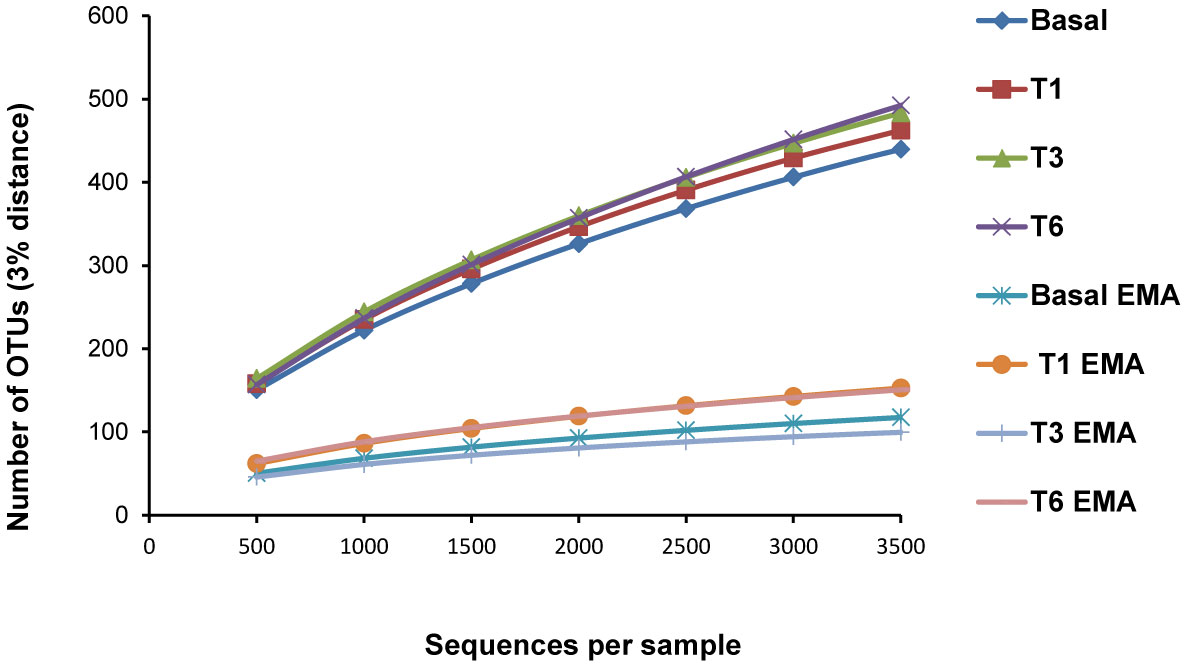
Figures(4) / Tables(2)

Lucía Guadamuro, M. Andrea Azcárate-Peril, Rafael Tojo, Baltasar Mayo, Susana Delgado. Use of high throughput amplicon sequencing and ethidium monoazide dye to track microbiota changes in an equol-producing menopausal woman receiving a long-term isoflavones treatment[J]. AIMS Microbiology, 2019, 5(1): 102-116. doi: 10.3934/microbiol.2019.1.102







 DownLoad:
DownLoad: