Citation: Maria C D’Ovidio, Annarita Wirz, Danila Zennaro, Stefania Massari, Paola Melis, Vittoria M Peri, Chiara Rafaiani, Maria C Riviello, Adriano Mari. Biological occupational allergy: Protein microarray for the study of laboratory animal allergy (LAA)[J]. AIMS Public Health, 2018, 5(4): 352-365. doi: 10.3934/publichealth.2018.4.352

| [1] |
Gautrin D, Newman-Taylor AJ, Nordman H, et al. (2003) Controversies in epidemiology of occupational asthma. Eur Respir J 22: 551–559. doi: 10.1183/09031936.03.00047803

|
| [2] |
Bush RK (2001) Assessment and treatment of laboratory animal allergy. ILAR J 42: 55–64. doi: 10.1093/ilar.42.1.55
|
| [3] | Vandenplas O, Worm M, Cullinan P, et al. (2013) Occupational allergy, In: Pawankar R, Canonica GW, Holgate ST, et al., White Book on Allergy: Update 2013, World Allergy Organization (WAO), 72–77. |
| [4] |
Patel O, Syamlal G, Wood J, et al. (2018) Asthma mortality among persons aged 15–64 years, by industry and occupation-United States, 1999–2016. MMWR Morb Mortal Wkly Rep 67: 60–65. doi: 10.15585/mmwr.mm6702a2
|
| [5] | National Institute for Occupational Safety and Health (NIOSH) (1998) Preventing asthma in animal handlers. Atlanta (GA): Centers for Disease Control and Prevention, 97–116. |
| [6] |
Bush R (2001) Mechanism and epidemiology of laboratory animal allergy. ILAR J 42: 4–11. doi: 10.1093/ilar.42.1.4
|
| [7] | D'Ovidio MC, Martini A, Melis P, et al. (2011) Value of the microarray for the study of laboratory animal allergy (LAA). G Ital Med Lav Ergon 33: 109–116. |
| [8] |
Bhabha FK, Nixon R (2012) Occupational exposure to laboratory animals causing a severe exacerbation of atopic eczema. Australas J Dermatol 53: 155–156. doi: 10.1111/j.1440-0960.2011.00754.x
|
| [9] |
Bush R, Wood R, Eggleston P (1998) Laboratory animal allergy. J Allergy Clin Immunol 102: 99–112. doi: 10.1016/S0091-6749(98)70060-0
|
| [10] |
Acton D, Mccauley L (2007) Laboratory animal allergy: An occupational hazard. AAOHN J 55: 241–244. doi: 10.1177/216507990705500604
|
| [11] | Seward JP (1999) Occupational allergy to animals. Occup Med 14: 285–302. |
| [12] |
Gordon S (2001) Laboratory animal allergy: A British perspective on a global problem. ILAR J 42: 37–46. doi: 10.1093/ilar.42.1.37
|
| [13] |
Gordon S, Preece R (2003) Prevention of laboratory animal allergy. Occup Med 53: 371–377. doi: 10.1093/occmed/kqg117
|
| [14] |
Elliott L, Heederik D, Marshall S, et al. (2005) Incidence of allergy and allergy symptoms among workers exposed to laboratory animals. Occup Environ Med 62: 766–771. doi: 10.1136/oem.2004.018739
|
| [15] |
Muzembo BA, Eitoku M, Inaoka Y, et al. (2014) Prevalence of occupational allergy in medical researchers exposed to laboratory animals. Ind Health 52: 256–261. doi: 10.2486/indhealth.2012-0155
|
| [16] |
Jones M (2015) Laboratory animal allergy in the modern era. Curr Allergy Asthma Rep 15: 73. doi: 10.1007/s11882-015-0575-4
|
| [17] |
Cullinan P, Lowson D, Nieuwenhuijsen MJ, et al. (1994) Work related symptoms, sensitization and estimated exposure in workers not previously exposed to laboratory rats. Occup Environ Med 51: 589–592. doi: 10.1136/oem.51.9.589
|
| [18] | Draper A, Newman TA, Cullinan P (2003) Estimating the incidence of occupational asthma and rhinitis from laboratory animal allergens in the UK, 1999–2000. Occup Environ Med 60: 604–605. |
| [19] |
Nieuwenhuijsen MJ, Putcha V, Gordon S, et al. (2003) Exposure-response relations among laboratory animal workers exposed to rats. Occup Environ Med 60: 104–108. doi: 10.1136/oem.60.2.104
|
| [20] | Gordon S, Newman-Taylor AJ (1999) Animal, insect and shellfish allergy, In: Bernstein IL, Chan-Yeung M, Malo JL, et al. (eds), Asthma in the workplace, 2 Eds., New York: Marcel Dekker Inc, 399–424. |
| [21] |
Harrison DJ (2001) Controlling exposure to laboratory animal allergens. ILAR J 42: 17–35. doi: 10.1093/ilar.42.1.17
|
| [22] | European Union: European Commission, Commission Staff Working Document: Accompanying document to the Report from the Commission to the Council and the European Parliament. Seventh Report on the Statistics on the Number of Animals used for Experimental and other Scientific Purposes in the Member States of the European Union. 5 December 2013, COM (2013) 859 final, Available from: http://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:52013DC0859 [accessed 8 March 2018]. |
| [23] |
Longbottom JL, Price JA (1987) Allergy to laboratory animals: Characterization and source of two major mouse allergens, Ag 1 and Ag 3. Int Arch Allergy Immunol 82: 450–452. doi: 10.1159/000234251
|
| [24] |
Baker J, Berry A, Boscato LM, et al. (2001) Identification of some rabbit allergens as lipocalins. Clin Exp Allergy 31: 303–312. doi: 10.1046/j.1365-2222.2001.00973.x
|
| [25] |
Wood RA (2001) Laboratory animal allergens. ILAR J 42: 12–16. doi: 10.1093/ilar.42.1.12
|
| [26] |
Flower DR (1996) The lipocalin protein family: Structure and function. Biochem J 318: 1–14. doi: 10.1042/bj3180001
|
| [27] |
Mäntyjärvi R, Rautiainen J, Virtanen T (2000) Lipocalins as allergens. Biochim Biophys Acta 1482: 308–317. doi: 10.1016/S0167-4838(00)00139-4
|
| [28] |
Aalberse RC (2014) Mammalian airborne allergens. Chem Immunol Allergy 100: 243–247. doi: 10.1159/000358862
|
| [29] | Hunskaari S, Fosse RT (1990) Allergy to laboratory mice and rats: A review of the pathophysiology, epidemiology and clinical aspects. Lab Anim 24: 358–374. |
| [30] |
Siraganian R, Sandberg A (1979) Characterization of mouse allergens. J Allergy Clin Immunol 63: 435–442. doi: 10.1016/0091-6749(79)90219-7
|
| [31] |
Mari A, Ciardiello MA, Tamburrini M, et al. (2010) Proteomic analysis in the identification of allergenic molecules. Expert Rev Proteomics 7: 723–734. doi: 10.1586/epr.10.44
|
| [32] |
Bush RK, Stave GM (2003) Laboratory animal allergy: An update. ILAR J 44: 28–51. doi: 10.1093/ilar.44.1.28
|
| [33] |
Delclos GL, Arif AA, Aday L, et al. (2006) Validation of an asthma questionnaire for use in healthcare workers. Occup Environ Med 63: 173–179. doi: 10.1136/oem.2005.021634
|
| [34] |
Scala E, Alessandri C, Bernardi ML, et al. (2010) Cross-sectional survey on immunoglobulin E reactivity in 23,077 subjects using an allergenic molecule-based microarray detection system. Clin Exp Allergy 40: 911–921. doi: 10.1111/j.1365-2222.2010.03470.x
|
| [35] |
Mari A, Rasi C, Palazzo P, et al. (2009) Allergen databases: Current status and perspectives. Curr Allergy Asthma Rep 9: 376–383. doi: 10.1007/s11882-009-0055-9
|
| [36] |
Bernardi ML, Giangrieco I, Camardella L, et al. (2011) Allergenic lipid transfer proteins from plant-derived foods do not immunologically and clinically behave homogeneously: The kiwifruit LTP as a model. PLoS One 6: e27856. doi: 10.1371/journal.pone.0027856
|
| [37] | Liccardi G, Bilò MB, Manzi F, et al. (2015) What could be the role of molecular-based allergy diagnostics in detecting the risk of developing allergic sensitization to furry animals? Eur Ann Allergy Clin Immunol 47: 163–167. |
| [38] | Liccardi G, Meriggi A, Russo M, et al. (2015) The risk of sensitization to furry animals in patients already sensitized to cat/dog: An in vitro evaluation using molecular-based allergy diagnostics. J Allergy Clin Immunol 135: 1664–1666. |
| [39] | D'Ovidio MC, Martini A, Signorini S (2007) New tools for the study of occupational allergies related to animals. G Ital Med Lav Ergon 29: 489–491. |
| [40] |
Olivieri M, Biscardo CA, Palazzo P, et al. (2013) Wheat IgE profiling and wheat IgE levels in bakers with allergic occupational phenotypes. Occup Environ Med 70: 617–622. doi: 10.1136/oemed-2012-101112
|
| [41] |
van der WA, Lopata AL, Nieuwenhuizen NE, et al. (2010) Work-related allergy and asthma in spice mill workers-The impact of processing dried spices on IgE reactivity patterns. Int Arch Allergy Immunol 152: 271–278. doi: 10.1159/000283038
|
| [42] |
Feary J, Cullinan P (2016) Laboratory animal allergy: A new world. Curr Opin Allergy Clin Immunol 16: 107–112. doi: 10.1097/ACI.0000000000000256
|
| [43] |
Simoneti CS, Freitas AS, Barbosa MC, et al. (2016) Study of risk factors for atopic sensitization, asthma, and bronchial hyperresponsiveness in animal laboratory workers. J Occup Health 58: 7–15. doi: 10.1539/joh.15-0045-OA
|
| [44] |
Kampitak T, Betschel SD (2016) Anaphylaxis in laboratory workers because of rodent handling: Two case reports. J Occup Health 58: 381–383. doi: 10.1539/joh.16-0053-CS
|
| [45] | Italian Legislative Decree no. 81/2008. Implementation of the article 1 of the law 3 august 2007, no. 123 concerning the protection of health and safety in the workplaces. Ordinary Supplement no.108 of the Official Journal no. 101 of 30 April 2008. Available from: www.gazzettaufficiale.it/index.jsp. |
| [46] | Italian Legislative Decree no. 106/2009. Supplementary and corrective provisions of the legislative decree 9 april 2008, no. 81, concerning the protection of health and safety in the workplaces. Ordinary Supplement no. 142 of the Official Journal no. 180 of 05 August 2009. Available from: www.gazzettaufficiale.it/index.jsp. |
| [47] | Legislative Decree 4 march 2014, n. 26. Implementation of the Directive 2010/63/EU on the protection of animals used for scientific purposes (Official Journal no. 61 of 14 March 2014). Available from: www.gazzettaufficiale.it/index.jsp. |
| [48] | NRC-National Research Council (1997) Occupational health and safety in the care and use of research animals. Washington DC: National Academy Press, 1969: 51–64. |
| [49] | D'Ovidio MC, Wirz A, Grandi C, et al. Education and training in animal care: A workplace scenario to combine human health and animal health. Poster Section presented at: International Congress on Occupational Health. 31st ICOH; 2015 May 31–June 5; Seoul Korea. |
| [50] |
Stave GM (2018) Occupational animal allergy. Curr Allergy Asthma Rep 18: 11. doi: 10.1007/s11882-018-0755-0
|
| [51] |
Schelkle M, Braun J, Jörres R, et al. (2017) Respiratory allergies among veterinarians: Two cross-sectional surveys from 2006 to 2012. Int Arch Occup Environ Health 90: 639–643. doi: 10.1007/s00420-017-1226-y
|
| [52] | Lemaire M, Oppliger A, Hotz P, et al. (2017) Can serum cytokine profile discriminate irritant-induced and allergen-induced symptoms? A cross-sectional study in workers mostly exposed to laboratory animals. Occup Environ Med 74: 592–600. |
| [53] | Wirz A, D'Ovidio MC, Massari S, et al. (2016) Methodological approach for the study of LAA (Laboratory Animal Allergy): A proposal to harmonize the procedures of risk evaluation. XXII AISAL Symposium. Innovation in laboratory animals science. The future is now. 22nd–24th October 2015 Naples, Italy. Comp Med 66: 78. |
| [54] |
Westall L, Graham IR, Bussell J (2015) A risk-based approach to reducing exposure of staff to laboratory animal allergens. Lab Anim 44: 32–38. doi: 10.1038/laban.603
|
| [55] |
Mason HJ, Willerton L (2017) Airborne exposure to laboratory animal allergens. AIMS Allergy Immunol 1: 78–88. doi: 10.3934/Allergy.2017.2.78
|
| [56] |
Simoneti CS, Ferraz E, Menezes MBD, et al. (2017) Allergic sensitization to laboratory animals is more associated with asthma, rhinitis, and skin symptoms than sensitization to common allergens. Clin Exp Allergy 47: 1436–1444. doi: 10.1111/cea.12994
|
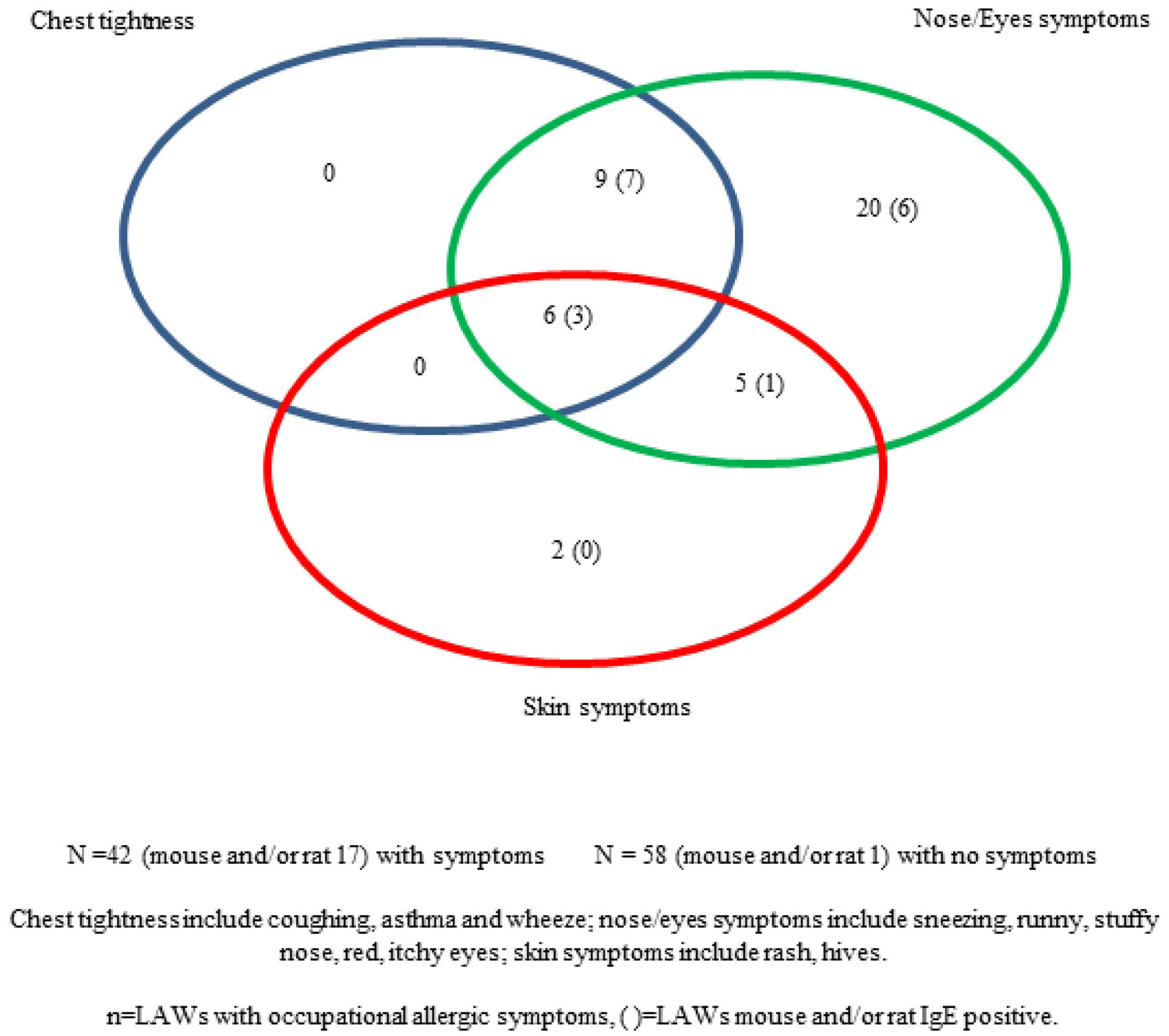

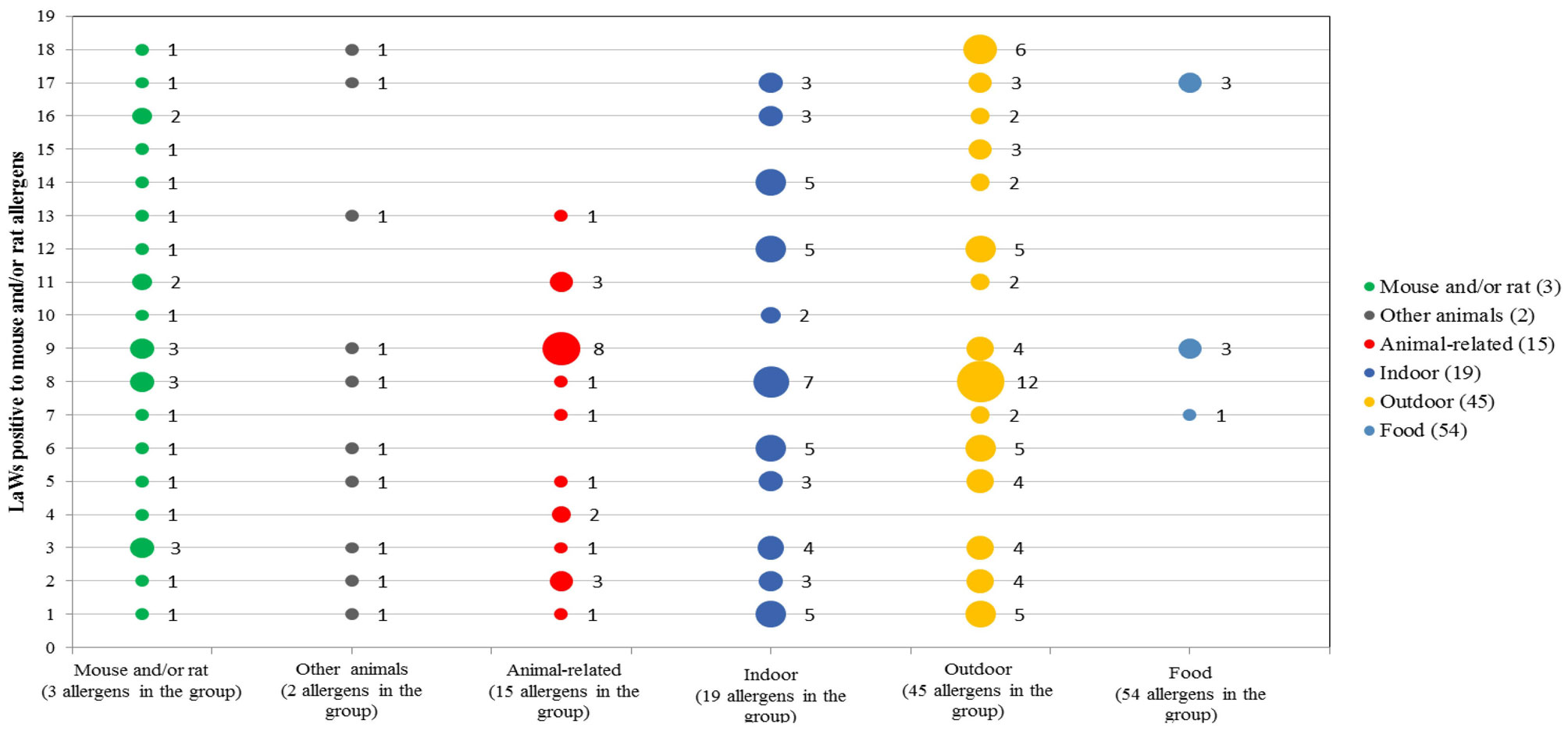
Figures(3) / Tables(3)

Maria C D’Ovidio, Annarita Wirz, Danila Zennaro, Stefania Massari, Paola Melis, Vittoria M Peri, Chiara Rafaiani, Maria C Riviello, Adriano Mari. Biological occupational allergy: Protein microarray for the study of laboratory animal allergy (LAA)[J]. AIMS Public Health, 2018, 5(4): 352-365. doi: 10.3934/publichealth.2018.4.352







 DownLoad:
DownLoad: