Citation: Hector Rudilla, Alexandra Merlos, Eulalia Sans-Serramitjana, Ester Fuste, Josep M. Sierra, Antonio Zalacain, Teresa Vinuesa, Miguel Vinas. New and old tools to evaluate new antimicrobial peptides[J]. AIMS Microbiology, 2018, 4(3): 522-540. doi: 10.3934/microbiol.2018.3.522

| [1] |
Donadio S, Maffioli S, Monciardini P, et al. (2010) Sources of novel antibiotics-aside the common roads. Appl Microbiol Biotechnol 88: 1261–1267. doi: 10.1007/s00253-010-2877-8

|
| [2] | Sengupta S, Chattopadhyay MK, Grossart HP (2013) The multifaceted roles of antibiotics and antibiotic resistance in nature. Front Microbiol 4: 47. |
| [3] |
Sans-Serramitjana E, Fusté E, Martínez-Garriga B, et al. (2016) Killing effect of nanoencapsulated colistin sulfate on Pseudomonas aeruginosa from cystic fibrosis patients. J Cyst Fibros 15: 611–618. doi: 10.1016/j.jcf.2015.12.005
|
| [4] |
Bilal M, Rasheed T, Iqbal HMN, et al. (2017) Macromolecular agents with antimicrobial potentialities: A drive to combat antimicrobial resistance. Int J Biol Macromol 103: 554–574. doi: 10.1016/j.ijbiomac.2017.05.071
|
| [5] |
Rabanal F, Grau-Campistany A, Vila-Farrés X, et al. (2015) A bioinspired peptide scaffold with high antibiotic activity and low in vivo toxicity. Sci Rep 5: 10558. doi: 10.1038/srep10558
|
| [6] |
Townsend CA (2016) Convergent biosynthetic pathways to β-lactam antibiotics. Curr Opin Chem Biol 35: 97–108. doi: 10.1016/j.cbpa.2016.09.013
|
| [7] |
Azmi F, Skwarczynski M, Toth I (2016) Towards the development of synthetic antibiotics: designs inspired by natural antimicrobial peptides. Curr Med Chem 23: 4610–4624. doi: 10.2174/0929867323666160825162435
|
| [8] | Oliver JD (2005) The viable but nonculturable state in bacteria. J Microbiol 43: 93–100. |
| [9] |
Rudilla H, Fusté E, Cajal Y, et al. (2016) Synergistic antipseudomonal effects of synthetic peptide AMP38 and carbapenems. Molecules 21: 1223. doi: 10.3390/molecules21091223
|
| [10] |
Jasim R, Schneider EK, Han M, et al. (2017) A fresh shine on cystic fibrosis inhalation therapy: Antimicrobial synergy of Polymyxin B in combination with silver nanoparticles. J Biomed Nanotechnol 13: 447–445. doi: 10.1166/jbn.2017.2355
|
| [11] |
Cavalcante CSP, de Aguiar FLL, Fontenelle ROS, et al. (2018) Insights into the candidacidal mechanism of Ctn[15-34]-a carboxyl-terminal, crotalicidin-derived peptide related to cathelicidins. J Med Microbiol 67: 129–138. doi: 10.1099/jmm.0.000652
|
| [12] |
Melikyan GB (2017) How entry inhibitors synergize to fight HIV. J Biol Chem 292: 16511–16512. doi: 10.1074/jbc.H117.791731
|
| [13] |
Betts J, Nagel C, Schatzschneider U, et al. (2017) Antimicrobial activity of carbon monoxide-releasing molecule [Mn(CO)3(tpa-κ3N)]Br versus multidrug-resistant isolates of Avian Pathogenic Escherichia coli and its synergy with colistin. PLoS One 12: e0186359. doi: 10.1371/journal.pone.0186359
|
| [14] |
Moon SH, Zhang X, Zheng G, et al. (2017) Novel linear lipopeptide paenipeptins with potential for eradicating biofilms and sensitizing Gram-negative bacteria to rifampicin and clarithromycin. J Med Chem 60: 9630–9640. doi: 10.1021/acs.jmedchem.7b01064
|
| [15] |
Hall-Stoodley L, Costerton JW, Stoodley P (2004) Bacterial biofilms: from the natural environment to infectious diseases. Nat Rev Microbiol 2: 95–108. doi: 10.1038/nrmicro821
|
| [16] |
Davies D (2003) Understanding biofilm resistance to antibacterial agents. Nat Rev Drug Discov 2: 114–122. doi: 10.1038/nrd1008
|
| [17] |
Döring G, Flume P, Heijerman H, et al. (2012) Treatment of lung infection in patients with cystic fibrosis: current and future strategies. J Cyst Fibros 11: 461–479. doi: 10.1016/j.jcf.2012.10.004
|
| [18] | Stewart PS (1996) Theoretical aspects of antibiotic diffusion into microbial biofilms. Antimicrob Agents Ch 40: 2517–2522. |
| [19] | Gilbert P, Maira-Litran T, McBain AJ, et al. (2002) The physiology and collective recalcitrance of microbial biofilm communities. Adv Microb Physiol 46: 202–256. |
| [20] |
Macià MD, Rojo-Molinero E, Oliver A (2014) Antimicrobial susceptibility testing in biofilm-growing bacteria. Clin Microbiol Infect 20: 981–990. doi: 10.1111/1469-0691.12651
|
| [21] | Neu TR, Lawrence JR (2014) Investigation of microbial biofilm structure by laser scanning microscopy. Adv Biochem Eng Biot 146: 1–51. |
| [22] |
Wright CJ, Shah MK, Powell LC, et al. (2010) Application of AFM from microbial cell to biofilm. Scanning 32: 134–149. doi: 10.1002/sca.20193
|
| [23] |
Mah TF, O'Toole GA (2001) Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol 9: 34–39. doi: 10.1016/S0966-842X(00)01913-2
|
| [24] | Ceri H, Olson ME, Stremick C, et al. (1999) The Calgary Biofilm Device: new technology for rapid determination of antibiotic susceptibilities of bacterial biofilms. J Clin Microbiol 37: 1771–1776. |
| [25] | Kostakioti M, Hadjifrangiskou M, Hultgren SJ (2013) Bacterial biofilms: development, dispersal, and therapeutic strategies in the dawn of the postantibiotic era. CSH Perspect Med 3: a010306. |
| [26] |
Toutain CM, Caizza NC, Zegans ME, et al. (2007) Roles for flagellar stators in biofilm formation by Pseudomonas aeruginosa. Res Microbiol 158: 471–477. doi: 10.1016/j.resmic.2007.04.001
|
| [27] |
Pratt LA, Kolter R (1998) Genetic analysis of Escherichia coli biofilm formation: roles of flagella, motility, chemotaxis and type I pili. Mol Microbiol 30: 285–293. doi: 10.1046/j.1365-2958.1998.01061.x
|
| [28] | Kwasny SM, Opperman TJ (2010) Static biofilm cultures of Gram-positive pathogens grown in a microtiter format used for anti-biofilm drug discovery. Curr Protoc Pharmacol 50: 13A.8.1–13A.8.23. |
| [29] |
Moskowitz SM, Foster JM, Emerson J, et al. (2004) Clinically feasible biofilm susceptibility assay for isolates of Pseudomonas aeruginosa from patients with cystic fibrosis. J Clin Microbiol 42: 1915–1922. doi: 10.1128/JCM.42.5.1915-1922.2004
|
| [30] |
Bhattacharya M, Wozniak DJ, Stoodley P, et al. (2015) Prevention and treatment of Staphylococcus aureus biofilms. Expert Rev Anti-Infe 13: 1499–1516. doi: 10.1586/14787210.2015.1100533
|
| [31] |
Francolini I, Donelli G (2010) Prevention and control of biofilm-based medical-device-related infections. FEMS Immunol Med Mic 59: 227–238. doi: 10.1111/j.1574-695X.2010.00665.x
|
| [32] | Doring G, Hoiby N (2004) Early intervention and prevention of lung disease in cystic fibrosis: a European consensus. J Cyst Fibros 3: 67–91. |
| [33] |
Fernández-Olmos A, García-Castillo M, Maiz L (2012) In vitro prevention of Pseudomonas aeruginosa early biofilm formation with antibiotics used in cystic fibrosis patients. Int J Antimicrob Ag 40: 173–176. doi: 10.1016/j.ijantimicag.2012.04.006
|
| [34] |
Sans-Serramitjana E, Jorba M, Fusté E, et al. (2017) Free and nanoencapsulated tobramycin: effects on planktonic and biofilm forms of Pseudomonas. Microorganisms 5: 35. doi: 10.3390/microorganisms5030035
|
| [35] | Zhou YF, Shi W, Yu Y, et al. (2015) Pharmacokinetic/Pharmacodynamic correlation of cefquinome against experimental catheter-associated biofilm infection due to Staphylococcus aureus. Front Microbiol 6: 1513. |
| [36] |
Moormeier DE, Bayles KW (2017) Staphylococcus aureus biofilm: a complex developmental organism. Mol Microbiol 104: 365–376. doi: 10.1111/mmi.13634
|
| [37] |
Sans-Serramitjana E, Jorba M, Pedraz JL, et al. (2017) Determination of the spatiotemporal dependence of Pseudomonas aeruginosa biofilm viability after treatment with NLC-colistin. Int J Nanomed 12: 4409–4413. doi: 10.2147/IJN.S138763
|
| [38] |
Quilès F, Saadi S, Francius G, et al. (2016) In situ and real time investigation of the evolution of a Pseudomonas fluorescens nascent biofilm in the presence of an antimicrobial peptide. BBA-Biomembranes 1858: 75–84. doi: 10.1016/j.bbamem.2015.10.015
|
| [39] |
Pamp SJ, Gjermansen M, Johansen HK, et al. (2008) Tolerance to the antimicrobial peptide colistin in Pseudomonas aeruginosa biofilms is linked to metabolically active cells, and depends on the pmr and mexAB-oprM genes. Mol Microbiol 68: 223–240. doi: 10.1111/j.1365-2958.2008.06152.x
|
| [40] |
Hengzhuang W, Ciofu O, Yang L, et al. (2013) High b-lactamase levels change the pharmacodynamics of b-lactam antibiotics in Pseudomonas aeruginosa biofilms. Antimicrob Agents Ch 57: 196–204. doi: 10.1128/AAC.01393-12
|
| [41] | Molecular probes, Product information LIVE/DEAD ® BacLight™ bacterial viability Kit, 2001. |
| [42] |
Binnig G, Quate CF, Gerber C (1986) Atomic force microscope. Phys Rev Lett 56: 930–933. doi: 10.1103/PhysRevLett.56.930
|
| [43] |
Li A, Ho B, Ding JL, et al. (2010) Use of atomic force microscopy as a tool to understand the action of antimicrobial peptides on bacteria. Method Mol Biol 618: 235–247. doi: 10.1007/978-1-60761-594-1_15
|
| [44] | López-Jiménez L, Arnabat-Domínguez J, Viñas M, et al. (2015) Atomic force microscopy visualization of injuries in Enterococcus faecalis surface caused by Er,Cr:YSGG and diode lasers. Med Oral Patol Oral Cir Bucal 20: e45–e51. |
| [45] | Zalacain A, Merlos A, Planell E, et al. (2017) Clinical laser treatment of toenail onychomycoses. Laser Med Sci 33: 927–933. |
| [46] |
Gonçalves S, Silva PM, Felício MR, et al. (2017) Psd1 effects on Candida albicans planktonic cells and biofilms. Front Cell Infect Mi 7: 249. doi: 10.3389/fcimb.2017.00249
|
| [47] |
Mularski A, Wilksch JJ, Wang H, et al. (2015) Atomic force microscopy reveals the mechanobiology of lytic peptide action on bacteria. Langmuir 31: 6164–6171. doi: 10.1021/acs.langmuir.5b01011
|
| [48] | Ramalingam B, Parandhaman T, Das SK (2016) Antibacterial effects of biosynthesized silver nanoparticles on surface ultrastructure and nanomechanical properties of Gram-negative bacteria viz. Escherichia coli and Pseudomonas aeruginosa. ACS Appl Mater Inter 8: 4963–4976. |
| [49] |
Canetta E, Riches A, Borger E, et al. (2014) Discrimination of bladder cancer cells from normal urothelial cells with high specificity and sensitivity: combined application of atomic force microscopy and modulated Raman spectroscopy. Acta Biomater 10: 2043–2055. doi: 10.1016/j.actbio.2013.12.057
|
| [50] |
Henderson JM, Waring AJ, Separovic F, et al. (2016) Antimicrobial peptides share a common interaction driven by membrane line tension reduction. Biophys J 111: 2176–2189. doi: 10.1016/j.bpj.2016.10.003
|
| [51] |
Angélique L, Frederik W, Garmi J, et al. (2015) The potential use of natural and structural analogues of antimicrobial peptides in the fight against neglected tropical diseases. Molecules 20: 15392–15433. doi: 10.3390/molecules200815392
|
| [52] |
Vinuesa T, Herráez R, Oliver L, et al. (2017) Benznidazole nanoformulates: a chance to improve therapeutics for chagas disease. Am J Trop Med Hyg 97: 1469–1476. doi: 10.4269/ajtmh.17-0044
|
| [53] |
Luque-Ortega JR, Rivas L (2010) Characterization of the leishmanicidal activity of antimicrobial peptides. Method Mol Biol 618: 393–420. doi: 10.1007/978-1-60761-594-1_25
|
| [54] |
Pinto EG, Pimenta DC, Antoniazzi MM, et al. (2013) Antimicrobial peptides isolated from Phyllomedusa nordestina (Amphibia) alter the permeability of plasma membrane of Leishmania and Trypanosoma cruzi. Exp Parasitol 135: 655–660. doi: 10.1016/j.exppara.2013.09.016
|
| [55] |
Tejería A, Pérez-Pertejo Y, Reguera RM, et al. (2016) Antileishmanial effect of new indeno-1,5-naphthyridines, selective inhibitors of Leishmania infantum type IB DNA topoisomerase. Eur J Med Chem 124: 740–749. doi: 10.1016/j.ejmech.2016.09.017
|
| [56] | Malucelli MI, Niero R, Lucchiari PH, et al. (1995) Evaluation of the polarographic technique for assay of the viability of freeze-dried BCG vaccine: II. Viability of the vaccine assessed by polarography, Warburg respirometry and colony counting. Vaccine 13: 273–275. |
| [57] | Wayne PA (2002) Reference method for broth dilution antifungal susceptibility testing of yeasts. |
| [58] | John HR (2008) Reference method for broth dilution antifungal susceptibility testing of filamentous fungi. |
| [59] | EUCAST method for susceptibility testing of yeasts (version 7.3.1), 2017. Available from: http://www.eucast.org/ast_of_fungi/methodsinantifungalsusceptibilitytesting/susceptibility_testing_of_yeasts/. |
| [60] | EUCAST method for susceptibility testing of moulds (version 9.3.1), 2017. Available from: http://www.eucast.org/ast_of_fungi/methodsinantifungalsusceptibilitytesting/susceptibility_testing_of_moulds/. |
| [61] |
Simitsopoulou M, Chatzimoschou A, Roilides E (2016) Candida species: methods and protocols, methods in molecular biology. Method Mol Biol 1356: 183–197. doi: 10.1007/978-1-4939-3052-4_14
|
| [62] |
Vale-Silva LA, Buchta V (2006) Antifungal susceptibility testing by flow cytometry: is it the future? Mycoses 49: 261–273. doi: 10.1111/j.1439-0507.2006.01243.x
|
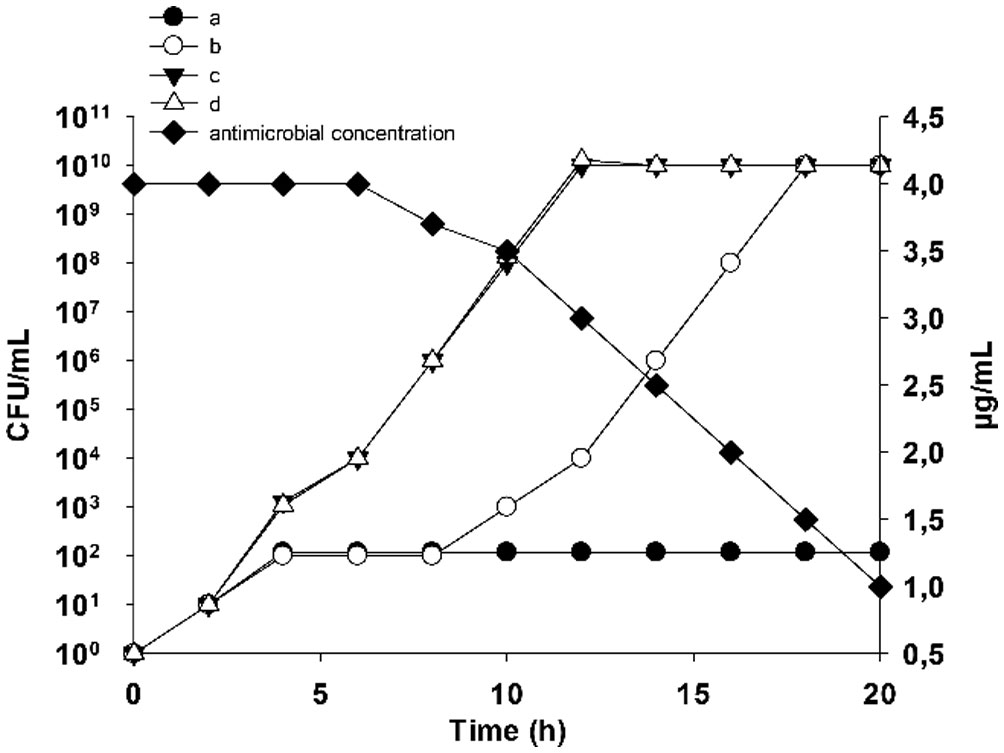
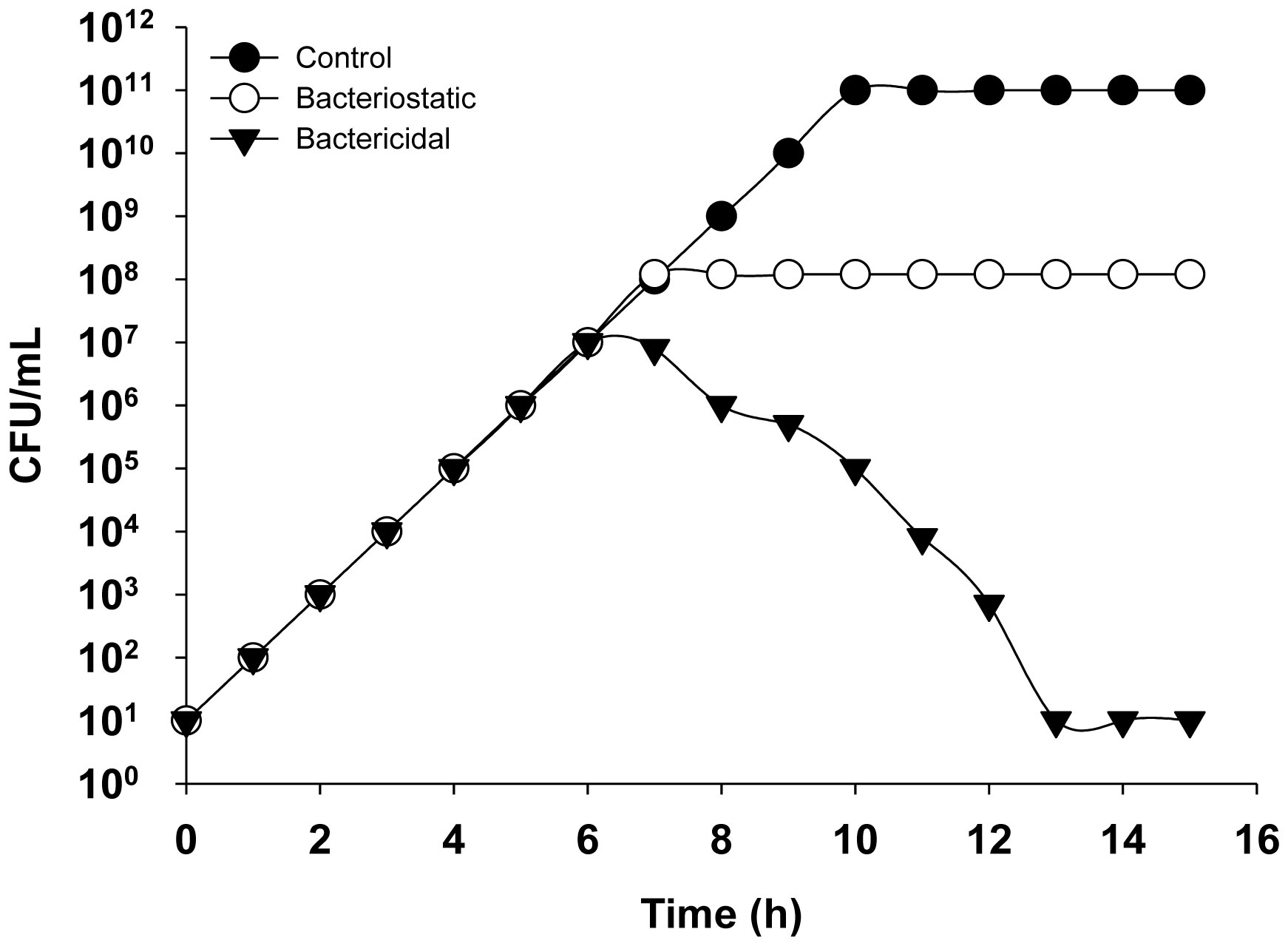
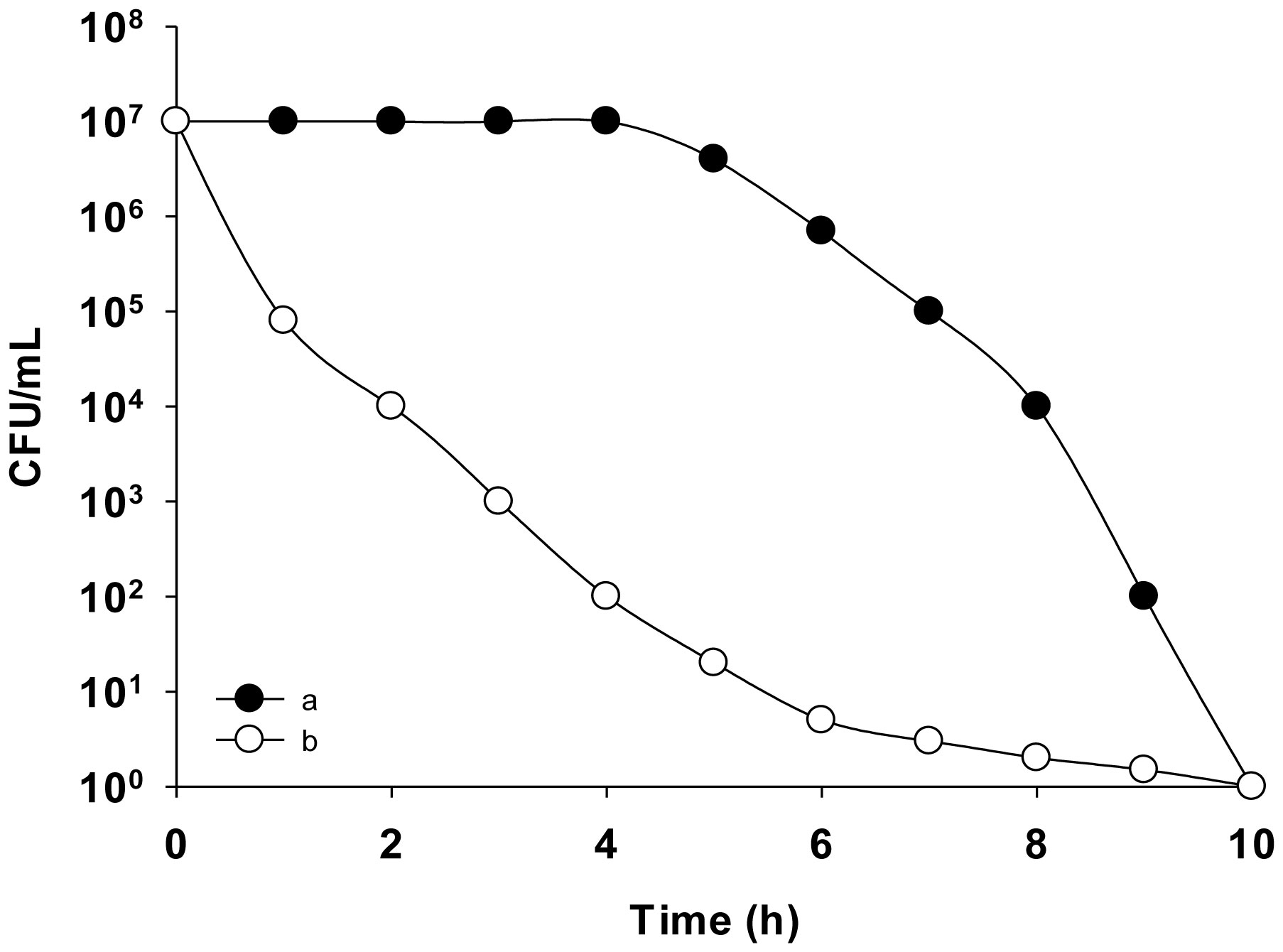
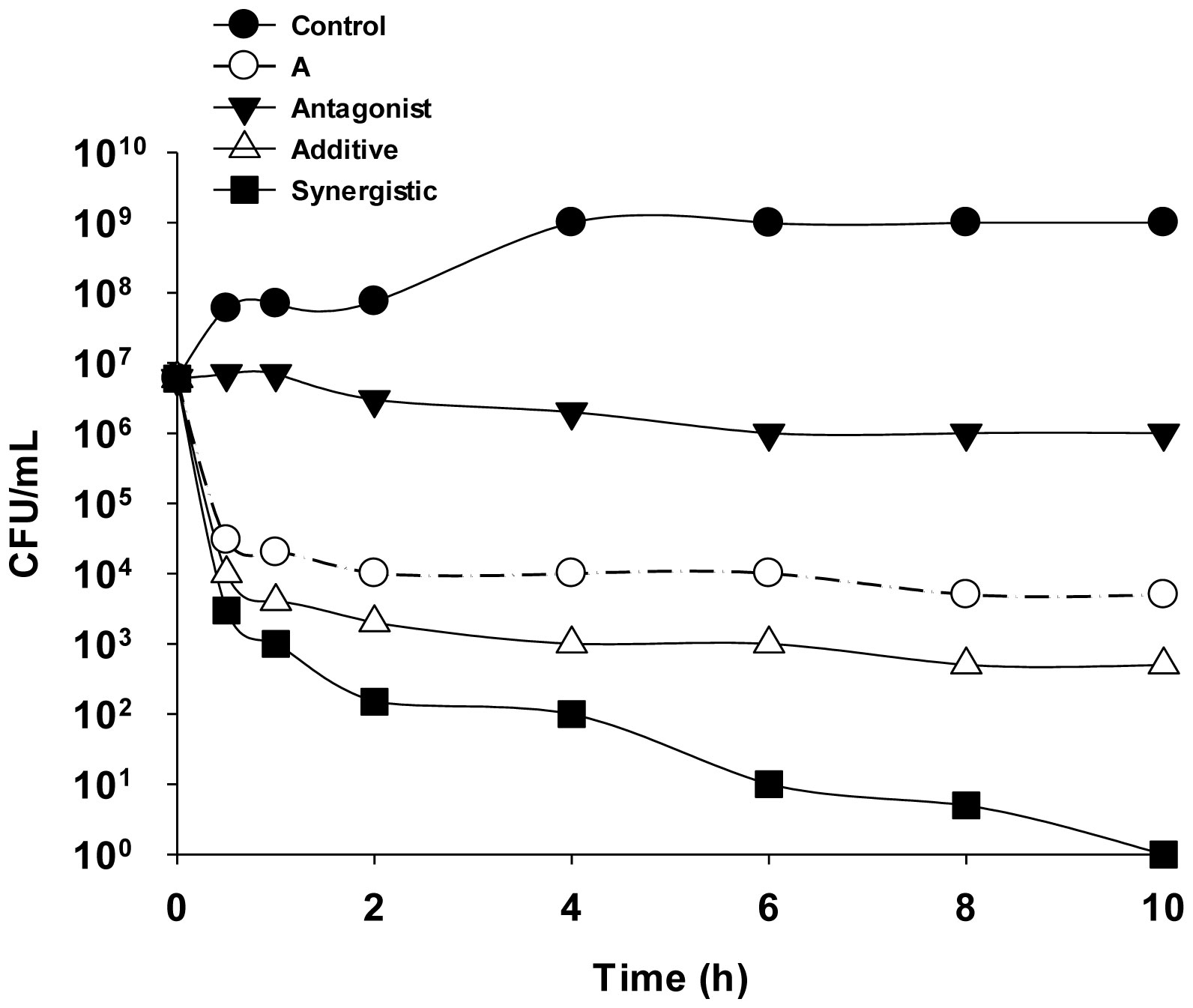
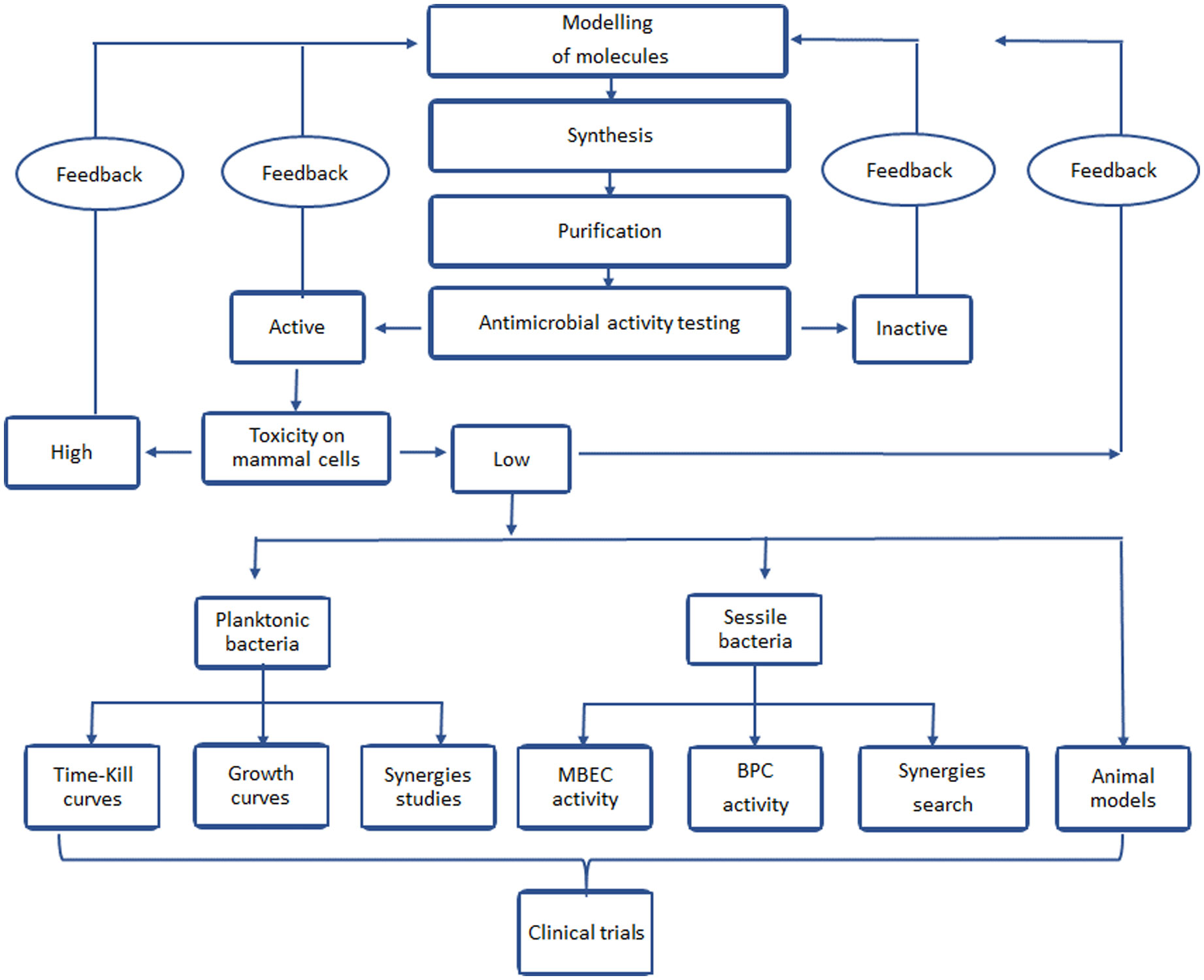
Figures(9)

Hector Rudilla, Alexandra Merlos, Eulalia Sans-Serramitjana, Ester Fuste, Josep M. Sierra, Antonio Zalacain, Teresa Vinuesa, Miguel Vinas. New and old tools to evaluate new antimicrobial peptides[J]. AIMS Microbiology, 2018, 4(3): 522-540. doi: 10.3934/microbiol.2018.3.522







 DownLoad:
DownLoad: