Citation: Fernanda Costa Brandão Berti, Karen Brajão de Oliveira. IL-10 in cancer: Just a classical immunosuppressive factor or also an immunostimulating one?[J]. AIMS Allergy and Immunology, 2018, 2(2): 88-97. doi: 10.3934/Allergy.2018.2.88

| [1] | Gopal M (2015) Role of cytokines in tumor immunity and immune tolerance to cancer, In: Rezaei N, Cancer Immunology: A Translational Medicine Context, 1 Ed., Heidelberg-New York-Dordrecht-London: Springer, 93–120. |
| [2] |
Grivennikov SI, Greten FR, Karin M (2010) Immunity, inflammation, and cancer. Cell 140: 883–899. doi: 10.1016/j.cell.2010.01.025

|
| [3] |
Mannino MH, Zhu Z, Xiao H, et al. (2015) The paradoxical role of IL-10 in immunity and cancer. Cancer Lett 367: 103–107. doi: 10.1016/j.canlet.2015.07.009
|
| [4] |
Mumm JB, Oft M (2013) Pegylated IL-10 induces cancer immunity: the surprising role of IL-10 as a potent inducer of IFN-γ-mediated CD8(+) T cell cytotoxicity. Bioessays 35: 623–631. doi: 10.1002/bies.201300004
|
| [5] |
Saraiva M, Christensen JR, Veldhoen M, et al. (2009) Interleukin-10 production by Th1 cells requires interleukin-12-induced STAT4 transcription factor and ERK MAP kinase activation by high antigen dose. Immunity 31: 209–219. doi: 10.1016/j.immuni.2009.05.012
|
| [6] | Trifunović J, Miller L, Debeljak Ž, et al. (2015) Pathologic patterns of interleukin 10 expression -a review. Biochem Med 25: 36–48. |
| [7] |
Sato T, Terai M, Tamura Y, et al. (2011) Interleukin 10 in the tumor microenvironment: A target for anticancer immunotherapy. Immunol Res 51: 170–182. doi: 10.1007/s12026-011-8262-6
|
| [8] |
Yoon BS, Kim YT, Kim JW, et al. (2007) Expression of human leukocyte antigen‑G and its correlation with interleukin‑10 expression in cervical carcinoma. Int J Gynecol Obstet 98: 48–53. doi: 10.1016/j.ijgo.2007.03.041
|
| [9] |
Cosman D, Müllberg J, Sutherland CL, et al. (2001) ULBPs, novel MHC class I-related molecules, bind to CMV glycoprotein UL16 and stimulate NK cytotoxicity through the NKG2D receptor. Immunity 14: 123–133. doi: 10.1016/S1074-7613(01)00095-4
|
| [10] |
Moretta L, Bottino C, Pende D, et al. (2006) Surface NK receptors and their ligands on tumor cells. Semin Immunol 18: 151–158. doi: 10.1016/j.smim.2006.03.002
|
| [11] |
Groh V, Wu J, Yee C, et al. (2002) Tumour-derived soluble MIC ligands impair expression of NKG2D and T-cell activation. Nature 419: 734–738. doi: 10.1038/nature01112
|
| [12] |
Gasser S, Orsulic S, Brown EJ, et al. (2005) The DNA damage pathway regulates innate immune system ligands of the NKG2D receptor. Nature 436: 1186–1190. doi: 10.1038/nature03884
|
| [13] |
Smyth MJ, Swann J, Cretney E, et al. (2005) NKG2D function protects the host from tumor initiation. J Exp Med 202: 583–588. doi: 10.1084/jem.20050994
|
| [14] |
Koebel CM, Vermi W, Swann JB, et al. (2007) Adaptive immunity maintains occult cancer in an equilibrium state. Nature 450: 903–907. doi: 10.1038/nature06309
|
| [15] |
Urosevic M, Dummer R (2003) HLA-G and IL-10 expression in human cancer-different stories with the same message. Semin Cancer Biol 13: 337–342. doi: 10.1016/S1044-579X(03)00024-5
|
| [16] | Szekeres-Bartho J (2002) Immunological relationship between the mother and the fetus. Int Rev Immunol 6: 471–495. |
| [17] | Noguchi K, Isogai M, Kuwada E, et al. (2004) Detection of anti-HLA-F antibodies in sera from cancer patients. Anticancer Res 24: 3387–3392. |
| [18] |
Curigliano G, Criscitiello C, Gelao L, et al. (2013) Molecular pathways: human leukocyte antigen G (HLA-G). Clin Cancer Res 19: 5564–5571. doi: 10.1158/1078-0432.CCR-12-3697
|
| [19] | Lin A, Yan WH (2015) HLA-G expression in cancers: roles in immune evasion, metastasis and target for therapy. Mol Med 21: 782–791. |
| [20] | Riteau B, Rouas-Freiss N, Menier C, et al. (2001) HLA-G2, -G3, and -G4 isoforms expressed as nonmature cell surface glycoproteins inhibit NK and antigen-specific CTL cytolysis. J Immunol 8: 5018–5026. |
| [21] |
Seo N, Hayakawa S, Tokura Y (2002) Mechanisms of immune privilege for tumor cells by regulatory cytokines produced by innate and acquired immune cells. Semin Cancer Biol 12: 291–300. doi: 10.1016/S1044-579X(02)00015-9
|
| [22] |
Roncarolo MG, Gregori S, Battaglia M, et al. (2006) Interleukin‑10‑secreting type 1 regulatory T cells in rodents and humans. Immunol Rev 212: 28–50. doi: 10.1111/j.0105-2896.2006.00420.x
|
| [23] | Achyut BR, Arbab AS (2016) Myeloid cell signatures in tumor microenvironment predicts therapeutic response in cancer. Oncotargets Ther 1: 1047–1055. |
| [24] |
Mocellin S, Marincola FM, Young HA (2005) Interleukin-10 and the immune response against cancer: a counterpoint. J Leukocyte Biol 78: 1043–1051. doi: 10.1189/jlb.0705358
|
| [25] | Lauw FN, Pajkrt D, Hack CE, et al. (2000) Proinflammatory effects of IL-10 during human endotoxemia. J Immunol 5: 2783–2789. |
| [26] | Kobayashi T, Matsuoka K, Sheikh SZ, et al. (2011) NFIL3 is a regulator of IL-12 p40 in macrophages and mucosal immunity. J Immunol 8: 4649–4655. |
| [27] | Mumm JB, Emmerich J, Zhang X, et al. (2011) IL-10 elicits IFNγ-dependent tumor immune surveillance. Cancer Cell 6: 781–796. |
| [28] |
Emmerich J, Mumm JB, Chan IH, et al. (2012) IL-10 directly activates and expands tumor-resident CD8(+) T cells without de novo infiltration from secondary lymphoid organs. Cancer Res 72: 3570–3581. doi: 10.1158/0008-5472.CAN-12-0721
|
| [29] |
Parato KG, Kumar A, Badley AD, et al. (2002) Normalization of natural killer cell function and phenotype with effective anti-HIV therapy and the role of IL-10. Aids 16: 1251–1256. doi: 10.1097/00002030-200206140-00007
|
| [30] | Mocellin S, Panelli MC, Wang E, et al. (2003) The dual role of IL-10. Trends Immunol 1: 36–43. |
| [31] | Mantovani A, Sozzani S, Locati M, et al. (2002) Macrophage polarization: tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol 11: 549–555. |
| [32] | Janjic BM, Lu G, Pimenov A, et al. (2002) Innate direct anticancer effector function of human immature dendritic cells. I. Involvement of an apoptosis-inducing pathway. J Immunol 4: 1823–1830. |
| [33] | Kelly JM, Darcy PK, Markby JL, et al. (2002) Induction of tumor-specific T cell memory by NK cell-mediated tumor rejection. Nat Immunol 1: 83–90. |
| [34] | Belardelli F, Ferrantini M (2002) Cytokines as a link between innate and adaptive antitumor immunity. Trends Immunol 4: 201–208. |
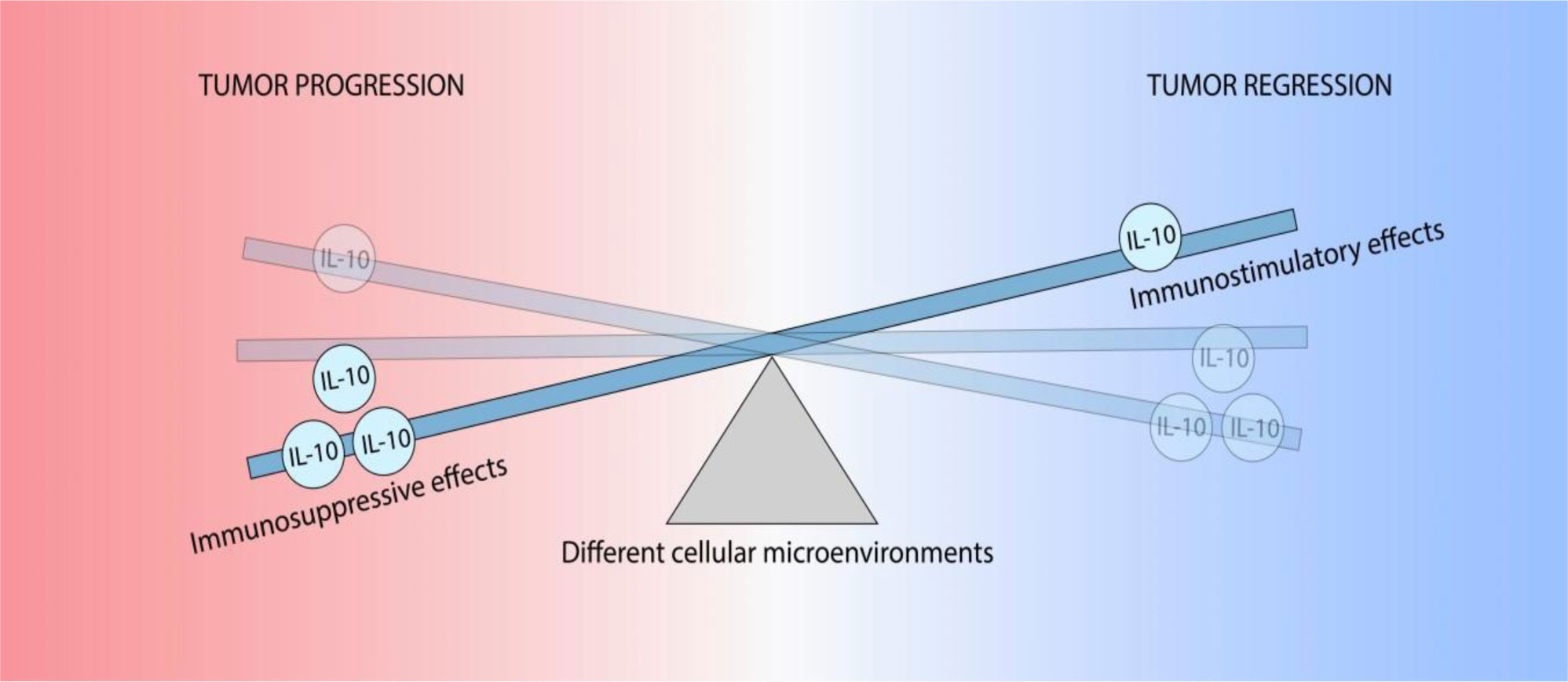
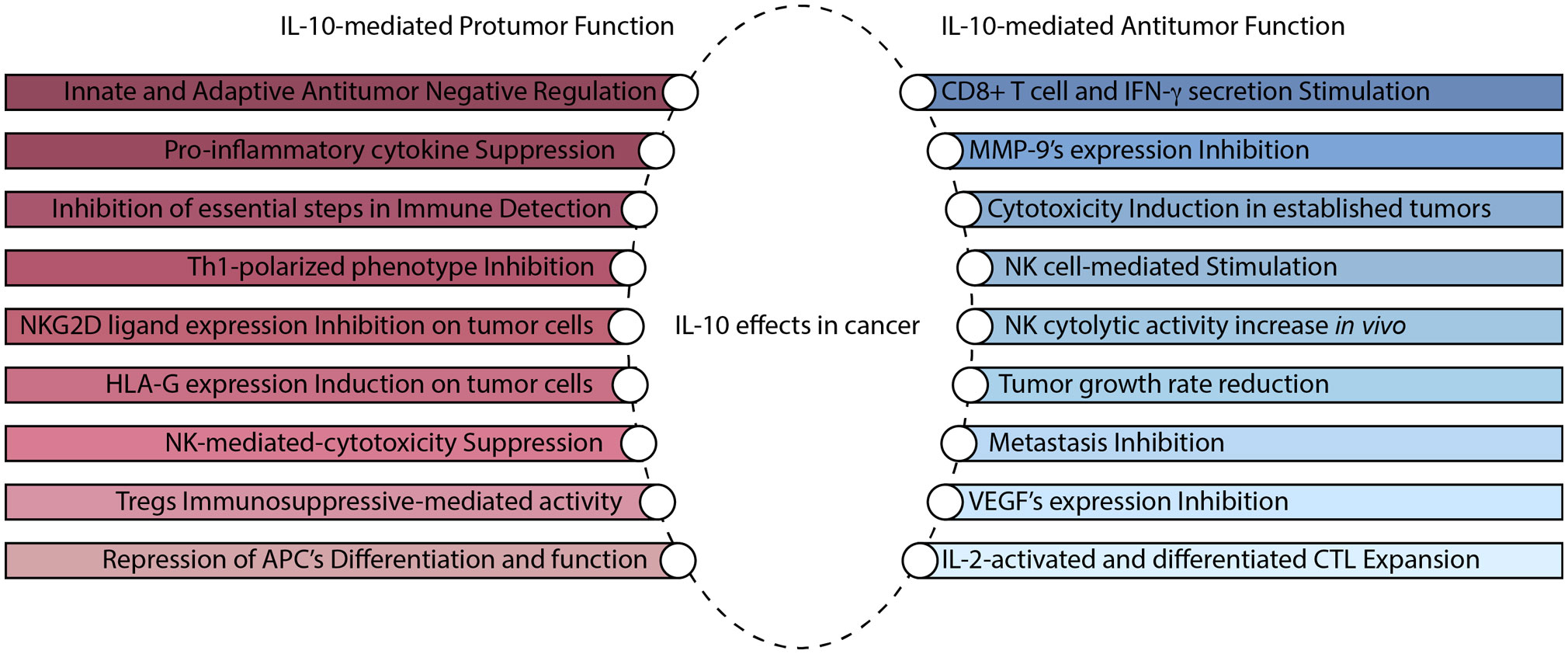
Figures(2)

Fernanda Costa Brandão Berti, Karen Brajão de Oliveira. IL-10 in cancer: Just a classical immunosuppressive factor or also an immunostimulating one?[J]. AIMS Allergy and Immunology, 2018, 2(2): 88-97. doi: 10.3934/Allergy.2018.2.88







 DownLoad:
DownLoad: