Citation: Hyunok Choi, Mark T. McAuley, David A. Lawrence. Prenatal exposures and exposomics of asthma[J]. AIMS Environmental Science, 2015, 2(1): 87-109. doi: 10.3934/environsci.2015.1.87

| [1] |
Adamko DJ, Sykes BD, Rowe BH (2012) The metabolomics of asthma: Novel diagnostic potential. Chest 141: 1295-1302. doi: 10.1378/chest.11-2028

|
| [2] | Wright RJ (2008) Stress and childhood asthma risk: overlapping evidence from animal studies and epidemiologic research. Allergy Asthma Clin Immun 4: 29. |
| [3] | Committee on the Assessment of Asthma and Indoor Air (2000) Clearing the Air:Asthma and Indoor Air Exposures; Division of Health Promotion DP, Institute of Medicine, editor: The National Academies Press. |
| [4] |
Asher MI, Montefort S, Björkstén B, et al. (2006) Worldwide time trends in the prevalence of symptoms of asthma, allergic rhinoconjunctivitis, and eczema in childhood: ISAAC Phases One and Three repeat multicountry cross-sectional surveys. Lancet 368: 733-743. doi: 10.1016/S0140-6736(06)69283-0
|
| [5] |
Asher M, Keil U, Anderson H, et al. (1995) International Study of Asthma and Allergies in Childhood (ISAAC): rationale and methods. Eur Respir J 8: 483-491. doi: 10.1183/09031936.95.08030483
|
| [6] |
Brim SN, Rudd RA, Funk RH, et al. (2008) Asthma Prevalence Among US Children in Underrepresented Minority Populations: American Indian/Alaska Native, Chinese, Filipino, and Asian Indian. Pediatrics 122: e217-e222. doi: 10.1542/peds.2007-3825
|
| [7] |
Kamble S, Bharmal M (2009) Incremental direct expenditure of treating asthma in the United States. J Asthma 46: 73-80. doi: 10.1080/02770900802503107
|
| [8] | Akinbami OJ, Moorman JE, Liu X (2011) Asthma prevalence, health care use, and mortality: United States, 2005-2009: US Department of Health and Human Services, Centers for Disease Control and Prevention, National Center for Health Statistics. |
| [9] |
Bryant-Stephens T (2009) Asthma disparities in urban environments. J Allergy Clinl Immunol 123: 1199-1206; quiz 1207-1198. doi: 10.1016/j.jaci.2009.04.030
|
| [10] |
Torgerson DG, Ampleford EJ, Chiu GY, et al. (2011) Meta-analysis of genome-wide association studies of asthma in ethnically diverse North American populations. Nat Genet 43: 887-892. doi: 10.1038/ng.888
|
| [11] |
Heinrich J (2011) Influence of indoor factors in dwellings on the development of childhood asthma. Int J Hyg Envir Heal 214: 1-25. doi: 10.1016/j.ijheh.2010.08.009
|
| [12] |
Busse PJ, Wang JJ, Halm EA (2005) Allergen sensitization evaluation and allergen avoidance education in an inner-city adult cohort with persistent asthma. J Allergy Clinl Immunol 116: 146-152. doi: 10.1016/j.jaci.2005.03.031
|
| [13] |
Jaakkola JJ, Jaakkola MS (2006) Professional cleaning and asthma. Curr Opin Allergy Clin Immunol 6: 85-90. doi: 10.1097/01.all.0000216849.64828.55
|
| [14] | Jaakkola MS, Jaakkola JJK (2004) Indoor Molds and Asthma in Adults. Advances in Applied Microbiology: Academic Press. pp. 309-338. |
| [15] |
Masoli M, Fabian D, Holt S, et al. (2004) Review article The global burden of asthma: executive summary of the GINA Dissemination Committee Report. Allergy 59: 469-478. doi: 10.1111/j.1398-9995.2004.00526.x
|
| [16] |
Yang IV, Schwartz DA (2012) Epigenetic mechanisms and the development of asthma. J Allergy Clin Immun 130: 1243-1255. doi: 10.1016/j.jaci.2012.07.052
|
| [17] |
Borish L, Culp JA (2008) Asthma: a syndrome composed of heterogeneous diseases. Ann Allergy Asthma Immunology Today 101: 1-8. doi: 10.1016/S1081-1206(10)60826-5
|
| [18] |
Ho S-M (2010) Environmental epigenetics of asthma: an update. J Allergy Clinl Immunol 126: 453-465. doi: 10.1016/j.jaci.2010.07.030
|
| [19] |
Ege MJ, Mayer M, Normand A-C, et al. (2011) Exposure to environmental microorganisms and childhood asthma. New Engl J Med 364: 701-709. doi: 10.1056/NEJMoa1007302
|
| [20] |
Hansel TT, Johnston SL, Openshaw PJ (2013) Microbes and mucosal immune responses in asthma. Lancet 381: 861-873. doi: 10.1016/S0140-6736(12)62202-8
|
| [21] |
Wild CP (2005) Complementing the genome with an "exposome": the outstanding challenge of environmental exposure measurement in molecular epidemiology. Cancer Epidem Biomar 14: 1847-1850. doi: 10.1158/1055-9965.EPI-05-0456
|
| [22] |
Larsson M, Weiss B, Janson S, et al. (2009) Associations between indoor environmental factors and parental-reported autistic spectrum disorders in children 6-8 years of age. Neurotoxicology 30: 822-831. doi: 10.1016/j.neuro.2009.01.011
|
| [23] | Bornehag CG, Sundell J, Sigsgaard T (2004) Dampness in buildings and health (DBH). Report from an on-going epidemiological investigation on the association between indoor environmental factors and health effects among children in Sweden. Indoor Air 14: 59-66. |
| [24] | Mendell MJ, Mirer AG, Cheung K, et al. (2011) Respiratory and Allergic Health Effects of Dampness, Mold, and Dampness-Related Agents: A Review of the Epidemiologic Evidence. Environ Health Perspect 119. |
| [25] |
Mendell MJ (2007) Indoor residential chemical emissions as risk factors for respiratory and allergic effects in children: a review. Indoor Air 17: 259-277. doi: 10.1111/j.1600-0668.2007.00478.x
|
| [26] |
Fang L, Clausen G, Fanger PO (1998) Impact of temperature and humidity on the perception of indoor air quality. Indoor air 8: 80-90. doi: 10.1111/j.1600-0668.1998.t01-2-00003.x
|
| [27] | Rosenbaum PF, Crawford JA, Anagnost SE, et al. (2009) Indoor airborne fungi and wheeze in the first year of life among a cohort of infants at risk for asthma. J Expos Sci Environ Epidemiol 20: 503-515. |
| [28] | Eurostat - The Statistical Office of the European Union (2010) Europe in Figures - EUROSTAT yearbook 2010. pp. Chapter 6. Living Conditions and Welfare. |
| [29] |
Samet JM, Spengler JD (2003) Indoor environments and health: moving into the 21st century. Am J Public Health 93: 1489-1493. doi: 10.2105/AJPH.93.9.1489
|
| [30] | Institute of Medicine (2004) Damp Indoor Spaces and Health. New York: The National Academies. |
| [31] |
Weschler CJ (2009) Changes in indoor pollutants since the 1950s. Atmos Environ 43: 153-169. doi: 10.1016/j.atmosenv.2008.09.044
|
| [32] |
Bönisch U, Böhme A, Kohajda T, et al. (2012) Volatile Organic Compounds Enhance Allergic Airway Inflammation in an Experimental Mouse Model. PLoS ONE 7: e39817. doi: 10.1371/journal.pone.0039817
|
| [33] |
Bornehag CG, Sundell J, Weschler CJ, et al. (2004) The association between asthma and allergic symptoms in children and phthalates in house dust: a nested case-control study. Environ health perspect 112: 1393-1397. doi: 10.1289/ehp.7187
|
| [34] | Herberth G, Herzog T, Hinz D, et al. (2012) Renovation activities during pregnancy induce a Th2 shift in fetal but not in maternal immune system. Int J Hyg Environ Health. |
| [35] | Bornehag CG, Sundell J, Hagerhed-Engman L, et al. (2005) "Dampness" at home and its association with airway, nose and skin symptoms among 10 851 preschool children in Sweden: a cross sectional study. Indoor Air 15: 48-55. |
| [36] |
Kim JL, Elfman L, Mi Y, et al. (2007) Indoor molds, bacteria, microbial volatile organic compounds and plasticizers in schools--associations with asthma and respiratory symptoms in pupils. Indoor Air 17: 153-163. doi: 10.1111/j.1600-0668.2006.00466.x
|
| [37] |
Wieslander G, Norback D (2010) Ocular symptoms, tear film stability, nasal patency, and biomarkers in nasal lavage in indoor painters in relation to emissions from water-based paint. Int Arch Occup Environ Health 83: 733-741. doi: 10.1007/s00420-010-0552-0
|
| [38] |
Wieslander G, Norback D (2010) A field study on clinical signs and symptoms in cleaners at floor polish removal and application in a Swedish hospital. Int Arch Occup Environ Health 83: 585-591. doi: 10.1007/s00420-010-0531-5
|
| [39] |
Wieslander G, Norback D, Edling C (1994) Occupational exposure to water based paint and symptoms from the skin and eyes. Occup Environ Med 51: 181-186. doi: 10.1136/oem.51.3.181
|
| [40] |
Wieslander G, Norback D, Edling C (1997) Airway symptoms among house painters in relation to exposure to volatile organic compounds (VOCS)--a longitudinal study. Ann Occup Hyg 41: 155-166. doi: 10.1093/annhyg/41.2.155
|
| [41] |
Wieslander G, Norback D, Nordstrom K, et al. (1999) Nasal and ocular symptoms, tear film stability and biomarkers in nasal lavage, in relation to building-dampness and building design in hospitals. Int Arch Occup Environ Health 72: 451-461. doi: 10.1007/s004200050398
|
| [42] |
Wieslander G, Kumlin A, Norback D (2010) Dampness and 2-ethyl-1-hexanol in floor construction of rehabilitation center: Health effects in staff. Arch Environ Occup Health 65: 3-11. doi: 10.1080/19338240903390248
|
| [43] |
Wieslander G, Lindgren T, Norback D, et al. (2000) Changes in the ocular and nasal signs and symptoms of aircrews in relation to the ban on smoking on intercontinental flights. Scand J Work Environ Health 26: 514-522. doi: 10.5271/sjweh.576
|
| [44] |
Wolkoff P, Schneider T, Kildesø J, et al. (1998) Risk in cleaning: chemical and physical exposure. Scie Total Environt 215: 135-156. doi: 10.1016/S0048-9697(98)00110-7
|
| [45] |
Zock JP, Plana E, Jarvis D, et al. (2007) The use of household cleaning sprays and adult asthma: an international longitudinal study. Am J Respir Crit Care Med 176: 735-741. doi: 10.1164/rccm.200612-1793OC
|
| [46] |
Choi H, Schmidbauer N, Sundell J, et al. (2010) Common household chemicals and the allergy risks in pre-school age children. PLoS One 5: e13423. doi: 10.1371/journal.pone.0013423
|
| [47] |
Ernstgård L, Lof A, Wieslander G, et al. (2007) Acute effects of some volatile organic compounds emitted from water-based paints. J Occup Environ Med 49: 880-889. doi: 10.1097/JOM.0b013e3181161ced
|
| [48] | IARC (2006) Formaldehyde, 2-Butoxyethanol and 1-tert-Butoxypropan-2-ol: Summary of Data Reported and Evaluation. In: ORGANIZATION WH, editor. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans. Lyon: WORLD HEALTH ORGANIZATION. |
| [49] | Garlantézec R, Warembourg C, Monfort C, et al. (2013) Urinary glycol ether metabolites in women and time to pregnancy: the PELAGIE cohort. Environ Health Perspect 121: 1167-1173. |
| [50] |
Korpi A, Järnberg J, Pasanen A-L (2009) Microbial volatile organic compounds. Crit rev toxicol 39: 139-193. doi: 10.1080/10408440802291497
|
| [51] | Korpi A, Pasanen A-L (1998) Volatile Compounds Originating for Mixed Microbial Cultures on Building Materials under various. Appl Environ Microb 64: 2914. |
| [52] | Norback D, Wieslander G, Nordstr K, et al. (2000) Asthma symptoms in relation to measured building dampness in upper concrete floor construction, and 2-ethyl-1-hexanol in indoor air. Int J Tuberc Lung D 4: 1016-1025. |
| [53] |
Nalli S, Horn OJ, Grochowalski AR, et al. (2006) Origin of 2-ethylhexanol as a VOC. Environ Pollut 140: 181-185. doi: 10.1016/j.envpol.2005.06.018
|
| [54] |
Goliff WS, Fitz DR, Cocker K, et al. (2012) Ambient measurements of 2,2,4-trimethyl, 1,3-pentanediol monoisobutyrate in Southern California. J Air Waste Manage 62: 680-685. doi: 10.1080/10962247.2012.666223
|
| [55] |
Järnström H, Saarela K, Kalliokoski P, et al. (2008) Comparison of VOC and ammonia emissions from individual PVC materials, adhesives and from complete structures. Environ Int 34: 420-427. doi: 10.1016/j.envint.2007.09.011
|
| [56] |
Maddalena R, Russell M, Sullivan DP, et al. (2009) Formaldehyde and Other Volatile Organic Chemical Emissions in Four FEMA Temporary Housing Units. Environmen Sci Technol 43: 5626-5632. doi: 10.1021/es9011178
|
| [57] | Wieslander G, Norback D, Bjornsson E, et al. (1997) Asthma and the indoor environment: the significance of emission of formaldehyde and volatile organic compounds from newly painted indoor surfaces. Int Arch Occup Environ Health 69: 115-124. |
| [58] |
Kempf M, Ramm S, Feuerbach T, et al. (2009) Occurrence of 2,2,4-trimethyl-1,3-pentanediol monoisobutyrate (Texanol®) in foods packed in polystyrene and polypropylene cups. Food Addit Contam A 26: 563-567. doi: 10.1080/02652030802562920
|
| [59] |
Sly PD, Boner AL, Björksten B, et al. (2008) Early identification of atopy in the prediction of persistent asthma in children. Lancet 372: 1100-1106. doi: 10.1016/S0140-6736(08)61451-8
|
| [60] |
Couraud S, Zalcman G, Milleron B, et al. (2012) Lung cancer in never smokers - A review. Eur J Cancer 48: 1299-1311. doi: 10.1016/j.ejca.2012.03.007
|
| [61] |
Wang L, Pinkerton KE (2007) Air pollutant effects on fetal and early postnatal development. Birth Defects Res Part C: Embryo Today: Rev 81: 144-154. doi: 10.1002/bdrc.20097
|
| [62] |
Wigle DT, Arbuckle TE, Walker M, et al. (2007) Environmental Hazards: Evidence for Effects on Child Health. J Toxicol Environmen Health B 10: 3-39. doi: 10.1080/10937400601034563
|
| [63] |
Burke H, Leonardi-Bee J, Hashim A, et al. (2012) Prenatal and Passive Smoke Exposure and Incidence of Asthma and Wheeze: Systematic Review and Meta-analysis. Pediatrics 129: 735-744. doi: 10.1542/peds.2011-2196
|
| [64] | Chambers DM, Ocariz JM, McGuirk MF, et al. (2011) Impact of cigarette smoking on Volatile Organic Compound (VOC) blood levels in the U.S. Population: NHANES 2003-2004. Environ Int 37: 1321-1328. |
| [65] | Gilbert CR, Arum SM, Smith CM (2009) Vitamin D deficiency and chronic lung disease. Can Respir J: Journal of the Canadian Thoracic Society 16: 75. |
| [66] |
Mansbach JM, Ginde AA, Camargo CA (2009) Serum 25-Hydroxyvitamin D Levels Among US Children Aged 1 to 11 Years: Do Children Need More Vitamin D? Pediatrics 124: 1404-1410. doi: 10.1542/peds.2008-2041
|
| [67] |
Zittermann A, Dembinski J, Stehle P (2004) Low vitamin D status is associated with low cord blood levels of the immunosuppressive cytokine interleukin-10. Pediatric Allergy Immunol 15: 242-246. doi: 10.1111/j.1399-3038.2004.00140.x
|
| [68] | Pfeffer PE, Hawrylowicz CM (2012) Vitamin D and lung disease. Thorax. |
| [69] |
Bozzetto S, Carraro S, Giordano G, et al. (2012) Asthma, allergy and respiratory infections: the vitamin D hypothesis. Allergy 67: 10-17. doi: 10.1111/j.1398-9995.2011.02711.x
|
| [70] |
Kozlowska E, Krzystyniak K, Drela N, et al. (1996) Thymus-directed immunotoxicity of airborne dust particles from Upper Silesia (Poland) under acute extrapulmonary studies in mice. J Toxicol Environmen Health 49: 563-579. doi: 10.1080/009841096160628
|
| [71] |
Busse W, Banks-Schlegel S, Noel P, et al. (2004) Future Research Directions in Asthma. Am J Resp Crit Care 170: 683-690. doi: 10.1164/rccm.200311-1539WS
|
| [72] | Holt PG, Macaubas C, Stumbles PA, et al. (1999) The role of allergy in the development of asthma. Nature 402: B12-17. |
| [73] |
Herberth G, Heinrich J, Röder S, et al. (2010) Reduced IFN-γ- and enhanced IL-4-producing CD4+ cord blood T cells are associated with a higher risk for atopic dermatitis during the first 2 yr of life. Pediatric Allergy Immunol 21: 5-13. doi: 10.1111/j.1399-3038.2009.00890.x
|
| [74] |
Herberth G, Hinz D, Roder S, et al. (2011) Maternal immune status in pregnancy is related to offspring's immune responses and atopy risk. Allergy 66: 1065-1074. doi: 10.1111/j.1398-9995.2011.02587.x
|
| [75] |
Lehmann I, Thoelke A, Rehwagen M, et al. (2002) The influence of maternal exposure to volatile organic compounds on the cytokine secretion profile of neonatal T cells. Environ Toxicol 17: 203-210. doi: 10.1002/tox.10055
|
| [76] |
Lehmann I, Thoelke A, Weiss M, et al. (2002) T cell reactivity in neonates from an East and a West German city - results of the LISA study. Allergy 57: 129-136. doi: 10.1046/j.0105-4538.2002.00001.x
|
| [77] |
Koike Y, Hisada T, Utsugi M, et al. (2007) Glutathione redox regulates airway hyperresponsiveness and airway inflammation in mice. Am J Respir Cell Mol Biol 37: 322-329. doi: 10.1165/rcmb.2006-0423OC
|
| [78] |
Kuipers H, Lambrecht BN (2004) The interplay of dendritic cells, Th2 cells and regulatory T cells in asthma. Curr Opin Immunol 16: 702-708. doi: 10.1016/j.coi.2004.09.010
|
| [79] | Holgate ST, Davies DE, Powell RM, et al. (2007) Local genetic and environmental factors in asthma disease pathogenesis: chronicity and persistence mechanisms. 29: 793-803. |
| [80] |
Ober C, Hoffjan S (2006) Asthma genetics 2006: the long and winding road to gene discovery. Genes Immun 7: 95-100. doi: 10.1038/sj.gene.6364284
|
| [81] |
Bilbo SD (2013) programming of neuroendocrine function by early-life experience: a critical role for the immune system. Horm Behav 63: 684-691. doi: 10.1016/j.yhbeh.2013.02.017
|
| [82] |
Mandal M, Donnelly R, Elkabes S, et al. (2013) Maternal immune stimulation during pregnancy shapes the immunological phenotype of offspring. Brain Behav Immun 33: 33-45. doi: 10.1016/j.bbi.2013.04.012
|
| [83] |
Bilbo SD, Schwarz JM (2012) The immune system and developmental programming of brain and behavior. Front Neuroendocrinol 33: 267-286. doi: 10.1016/j.yfrne.2012.08.006
|
| [84] |
Cory-Slechta DA, Virgolini MB, Rossi-George A, et al. (2008) Lifetime consequences of combined maternal lead and stress. Basic Clin Pharmacol Toxicol 102: 218-227. doi: 10.1111/j.1742-7843.2007.00189.x
|
| [85] |
Rappaport SM (2011) Implications of the exposome for exposure science. J Expo Sci Environ Epidemiol 21: 5-9. doi: 10.1038/jes.2010.50
|
| [86] |
Lewis R, Demmelmair H, Gaillard R, et al. (2013) The placental exposome: placental determinants of fetal adiposity and postnatal body composition. Ann Nutr Metab 63: 208-215. doi: 10.1159/000355222
|
| [87] | Miller GW, Jones DP (2013) The nature of nurture: refining the definition of the exposome. Toxicol Sci: kft251. |
| [88] |
Vineis P, Veldhoven K, Chadeau‐Hyam M, et al. (2013) Advancing the application of omics‐based biomarkers in environmental epidemiology. Environ Mol Mutagen 54: 461-467. doi: 10.1002/em.21764
|
| [89] |
Nicholson JK, Wilson ID (2003) Understanding'global'systems biology: metabonomics and the continuum of metabolism. Nat Rev Drug Discov 2: 668-676. doi: 10.1038/nrd1157
|
| [90] |
Thacher JD, Gruzieva O, Pershagen G, et al. (2014) Pre- and Postnatal Exposure to Parental Smoking and Allergic Disease Through Adolescence. Pediatrics 134: 428-434. doi: 10.1542/peds.2014-0427
|
| [91] |
Patel MM, Quinn JW, Jung KH, et al. (2011) Traffic density and stationary sources of air pollution associated with wheeze, asthma, and immunoglobulin E from birth to age 5 years among New York City children. Environ Res 111: 1222-1229. doi: 10.1016/j.envres.2011.08.004
|
| [92] |
Janssen B, Godderis L, Pieters N, et al. (2013) Placental DNA hypomethylation in association with particulate air pollution in early life. Part Fibre Toxicol 10: 22. doi: 10.1186/1743-8977-10-22
|
| [93] |
Janssen B, Munters E, Pieters N, et al. (2012) Placental Mitochondrial DNA Content and Particulate Air Pollution during in Utero Life. Environ Health Perspect 120: 1346-1352. doi: 10.1289/ehp.1104458
|
| [94] |
Lu L-JW, Anderson LM, Jones AB, et al. (1993) Persistence, gestation stage-dependent formation and interrelationship of benzo[a]pyrene-induced DNA adducts in mothers, placentae and fetuses of Erythrocebus patas monkeys. Carcinogenesis 14: 1805-1813. doi: 10.1093/carcin/14.9.1805
|
| [95] |
Bradding P, Walls AF, Holgate ST (2006) The role of the mast cell in the pathophysiology of asthma. J Allergy Clin Immunol 117: 1277-1284. doi: 10.1016/j.jaci.2006.02.039
|
| [96] |
Brightling CE, Bradding P, Symon FA, et al. (2002) Mast-cell infiltration of airway smooth muscle in asthma. N Engl J Med 346: 1699-1705. doi: 10.1056/NEJMoa012705
|
| [97] |
Peachell P (2005) Targeting the mast cell in asthma. Curr opin pharmacol 5: 251-256. doi: 10.1016/j.coph.2005.03.001
|
| [98] |
Kleeberger SR, Ohtsuka Y, Zhang L-Y, et al. (2001) Airway responses to chronic ozone exposure are partially mediated through mast cells. J Appl Physiol 90: 713-723. doi: 10.1063/1.1379355
|
| [99] |
Koren HS, Hatch GE, Graham DE (1990) Nasal lavage as a tool in assessing acute inflammation in response to inhaled pollutants. Toxicology 60: 15-25. doi: 10.1016/0300-483X(90)90159-E
|
| [100] |
Schierhorn K, Zhang M, Matthias C, et al. (1999) Influence of ozone and nitrogen dioxide on histamine and interleukin formation in a human nasal mucosa culture system. Am J Respir Cell Mol Biol 20: 1013-1019. doi: 10.1165/ajrcmb.20.5.3268
|
| [101] |
Shields RL, Gold WM (1987) Effect of inhaled ozone on lung histamine in conscious guinea pigs. Environ Res 42: 435-445. doi: 10.1016/S0013-9351(87)80211-6
|
| [102] |
Stenfors N, Pourazar J, Blomberg A, et al. (2002) Effect of ozone on bronchial mucosal inflammation in asthmatic and healthy subjects. Respir Med 96: 352-358. doi: 10.1053/rmed.2001.1265
|
| [103] |
Stenfors N, Bosson J, Helleday R, et al. (2010) Ozone exposure enhances mast-cell inflammation in asthmatic airways despite inhaled corticosteroid therapy. Inhal Toxicol 22: 133-139. doi: 10.3109/08958370903005736
|
| [104] |
Vagaggini B, Taccola M, Conti I, et al. (2001) Budesonide reduces neutrophilic but not functional airway response to ozone in mild asthmatics. Am J Resp Crit Care 164: 2172-2176. doi: 10.1164/ajrccm.164.12.2009090
|
| [105] |
Janssen BG, Munters E, Pieters N, et al. (2012) Placental mitochondrial DNA content and particulate air pollution during in utero life. Environ Health Perspect 120: 1346-1352. doi: 10.1289/ehp.1104458
|
| [106] |
Prahalad A, Manchester D, Hsu I, et al. (1999) Human placental microsomal activation and DNA adduction by air pollutants. B Environ Contam Tox 62: 93-100. doi: 10.1007/s001289900846
|
| [107] |
Rocha e Silva IR, Lichtenfels AJF, Amador Pereira LA, et al. (2008) Effects of ambient levels of air pollution generated by traffic on birth and placental weights in mice. Fertil Steril 90: 1921-1924. doi: 10.1016/j.fertnstert.2007.10.001
|
| [108] |
Topinka J, Binkova B, Mračková G, et al. (1997) DNA adducts in human placenta as related to air pollution and to GSTM1 genotype. Mutatio Res-Gen- Tox En 390: 59-68. doi: 10.1016/S0165-1218(96)00166-8
|
| [109] |
Veras MM, Damaceno-Rodrigues NR, Caldini EG, et al. (2008) Particulate urban air pollution affects the functional morphology of mouse placenta. Biol Reprod 79: 578-584. doi: 10.1095/biolreprod.108.069591
|
| [110] |
Fujimoto A, Tsukue N, Watanabe M, et al. (2005) Diesel exhaust affects immunological action in the placentas of mice. Environ Toxicol 20: 431-440. doi: 10.1002/tox.20129
|
| [111] | Menzies F, Shepherd M, Nibbs R, et al. (2010) The role of mast cells and their mediators in reproduction, pregnancy and labour. Hum reprod update: dmq053. |
| [112] | Woidacki K, Jensen F, Zenclussen AC (2013) Mast cells as novel mediators of reproductive processes. Front Immunol 4. |
| [113] | Szewczyk G, Pyzlak M, Klimkiewicz J, et al. (2012) Mast cells and histamine: do they influence placental vascular network and development in preeclampsia? Mediators Inflamm 2012. |
| [114] | Dadvand P, Figueras F, Basagana X, et al. (2013) Ambient air pollution and preeclampsia: a spatiotemporal analysis. Environ Health Perspect 121: 1365-1371. |
| [115] |
Lee PC, Roberts JM, Catov JM, et al. (2013) First trimester exposure to ambient air pollution, pregnancy complications and adverse birth outcomes in Allegheny County, PA. Matern Child Health J 17: 545-555. doi: 10.1007/s10995-012-1028-5
|
| [116] | Pereira G, Haggar F, Shand AW, et al. (2012) Association between pre-eclampsia and locally derived traffic-related air pollution: a retrospective cohort study. J Epidemiol Commun H |
| [117] |
Woo Y, Jeong D, Chung DH, et al. (2014) The Roles of Innate Lymphoid Cells in the Development of Asthma. Immune network 14: 171-181. doi: 10.4110/in.2014.14.4.171
|
| [118] | Vroman H, van den Blink B, Kool M (2014) Mode of dendritic cell activation; the decisive hand in Th2/Th17 cell differentiation. Implications in asthma severity? Immunobiology. |
| [119] |
Murata Y, Shimamura T, Hamuro J (2002) The polarization of Th1/Th2 balance is dependent on the intracellular thiol redox status of macrophages due to the distinctive cytokine production. Inter Immunol 14: 201-212. doi: 10.1093/intimm/14.2.201
|
| [120] |
Peterson JD, Herzenberg LA, Vasquez K, et al. (1998) Glutathione levels in antigen-presenting cells modulate Th1 versus Th2 response patterns. PNAS 95: 3071-3076. doi: 10.1073/pnas.95.6.3071
|
| [121] | Tuzova M, Jean J-C, Hughey RP, et al. (2014) Inhibiting lung lining fluid glutathione metabolism with GGsTop as a novel treatment for asthma. Front Pharmacol 5. |
| [122] |
Perzanowski MS, Miller RL, Tang D, et al. (2010) Prenatal acetaminophen exposure and risk of wheeze at age 5 years in an urban low-income cohort. Thorax 65: 118-123. doi: 10.1136/thx.2009.121459
|
| [123] | Penn AL, Rouse RL, Horohov DW, et al. (2007) In utero exposure to environmental tobacco smoke potentiates adult responses to allergen in BALB/c mice. Environ health perspect: 548-555. |
| [124] |
Raherison C, Pénard-Morand C, Moreau D, et al. (2007) In utero and childhood exposure to parental tobacco smoke, and allergies in schoolchildren. Resp med 101: 107-117. doi: 10.1016/j.rmed.2006.04.010
|
| [125] |
Jedrychowski WA, Perera FP, Majewska R, et al. (2014) Separate and joint effects of tranplacental and postnatal inhalatory exposure to polycyclic aromatic hydrocarbons: Prospective birth cohort study on wheezing events. Pediatr Pulmonol 49: 162-172. doi: 10.1002/ppul.22923
|
| [126] | Hansen S, Strøm M, Olsen SF, et al. (2013) Maternal concentrations of persistent organochlorine pollutants and the risk of asthma in offspring: results from a prospective cohort with 20 years of follow-up. |
| [127] | Whyatt RM, Perzanowski MS, Just AC, et al. (2014) Asthma in Inner-City Children at 5-11 Years of Age and Prenatal Exposure to Phthalates: The Columbia Center for Children’s Environmental Health Cohort. Environ Health Perspect. |
| [128] |
Spanier AJ, Kahn RS, Kunselman AR, et al. (2012) Prenatal exposure to bisphenol A and child wheeze from birth to 3 years of age. Environ health perspects 120: 916. doi: 10.1289/ehp.1104175
|
| [129] |
Stockinger B, Meglio PD, Gialitakis M, et al. (2014) The Aryl Hydrocarbon Receptor: Multitasking in the Immune System. Annu rev immunol 32: 403-432. doi: 10.1146/annurev-immunol-032713-120245
|
| [130] |
Krüger T, Long M, Bonefeld-Jørgensen EC (2008) Plastic components affect the activation of the aryl hydrocarbon and the androgen receptor. Toxicology 246: 112-123. doi: 10.1016/j.tox.2007.12.028
|
| [131] |
Cayrol C, Girard J-P (2014) IL-33: an alarmin cytokine with crucial roles in innate immunity, inflammation and allergy. Curr Opin Immunol 31: 31-37. doi: 10.1016/j.coi.2014.09.004
|
| [132] |
Allakhverdi Z, Comeau MR, Smith DE, et al. (2009) CD34+ hemopoietic progenitor cells are potent effectors of allergic inflammation. J Allergy Clinl Immunol 123: 472-478. e471. doi: 10.1016/j.jaci.2008.10.022
|
| [133] |
Préfontaine D, Lajoie-Kadoch S, Foley S, et al. (2009) Increased expression of IL-33 in severe asthma: evidence of expression by airway smooth muscle cells. J Immunol 183: 5094-5103. doi: 10.4049/jimmunol.0802387
|
| [134] |
Forsythe P, Ennis M (1999) Adenosine, mast cells and asthma. Inflamm Res 48: 301-307. doi: 10.1007/s000110050464
|
| [135] | Gao Y-d, Cao J, Li P, et al. (2014) Th2 cytokine-primed airway smooth muscle cells induce mast cell chemotaxis via secretion of ATP. J Asthma: 1-21. |
| [136] |
Mills KH, Dungan LS, Jones SA, et al. (2013) The role of inflammasome-derived IL-1 in driving IL-17 responses. J Leukoc Biol 93: 489-497. doi: 10.1189/jlb.1012543
|
| [137] |
Besnard A-G, Togbe D, Couillin I, et al. (2012) Inflammasome-IL-1-Th17 response in allergic lung inflammation. J Mol Cell Biol 4: 3-10. doi: 10.1093/jmcb/mjr042
|
| [138] |
Rappaport SM (2012) Biomarkers intersect with the exposome. Biomarkers 17: 483-489. doi: 10.3109/1354750X.2012.691553
|
| [139] |
Holmes E, Loo R, Stamler J, et al. (2008) Human metabolic phenotype diversity and its association with diet and blood pressure. Nature 453: 396 - 400. doi: 10.1038/nature06882
|
| [140] |
Tsai W, Chung R (2010) Viral hepatocarcinogenesis. Oncogene 29: 2309-2324. doi: 10.1038/onc.2010.36
|
| [141] |
Nicholson JK, Holmes E, Wilson ID (2005) Gut microorganisms, mammalian metabolism and personalized health care. Nat Rev Microb 3: 431-438. doi: 10.1038/nrmicro1152
|
| [142] |
Rappaport SM, Smith MT (2010) Environment and disease risks. Science(Washington) 330: 460-461. doi: 10.1126/science.1192603
|
| [143] |
Smith MT, Zhang L, McHale CM, et al. (2011) Benzene, the exposome and future investigations of leukemia etiology. Che Biol Interact 192: 155-159. doi: 10.1016/j.cbi.2011.02.010
|
| [144] |
Maitre L, Fthenou E, Athersuch T, et al. (2014) Urinary metabolic profiles in early pregnancy are associated with preterm birth and fetal growth restriction in the Rhea mother-child cohort study. BMC Medicine 12: 110. doi: 10.1186/1741-7015-12-110
|
| [145] |
Senn T, Hazen SL, Tang W (2012) Translating metabolomics to cardiovascular biomarkers. Prog Cardiovasc Dis 55: 70-76. doi: 10.1016/j.pcad.2012.06.004
|
| [146] |
Yang Y, Cruickshank C, Armstrong M, et al. (2013) New sample preparation approach for mass spectrometry-based profiling of plasma results in improved coverage of metabolome. J Chromatogr A 1300: 217-226. doi: 10.1016/j.chroma.2013.04.030
|
| [147] |
Kind T, Fiehn O (2010) Advances in structure elucidation of small molecules using mass spectrometry. Bioanalyt Rev 2: 23-60. doi: 10.1007/s12566-010-0015-9
|
| [148] |
Mattarucchi E, Baraldi E, Guillou C (2012) Metabolomics applied to urine samples in childhood asthma; differentiation between asthma phenotypes and identification of relevant metabolites. Biomed Chromatogr 26: 89-94. doi: 10.1002/bmc.1631
|
| [149] |
Ho WE, Xu Y-J, Xu F, et al. (2013) Metabolomics reveals altered metabolic pathways in experimental asthma. Am J Respir Cell Mol Biol 48: 204-211. doi: 10.1165/rcmb.2012-0246OC
|
| [150] |
Fanos V, Barberini L, Antonucci R, et al. (2011) Metabolomics in neonatology and pediatrics. Clin Biochem 44: 452-454. doi: 10.1016/j.clinbiochem.2011.03.006
|
| [151] |
Griffiths WJ, Koal T, Wang Y, et al. (2010) Targeted metabolomics for biomarker discovery. Angew Chem Int Edit 49: 5426-5445. doi: 10.1002/anie.200905579
|
| [152] |
Carraro S, Rezzi S, Reniero F, et al. (2007) Metabolomics applied to exhaled breath condensate in childhood asthma. Am J Resp Crit Care 175: 986-990. doi: 10.1164/rccm.200606-769OC
|
| [153] |
Tan YM, Conolly R, Chang DT, et al. (2012) Computational toxicology: application in environmental chemicals. Methods Mol Biol 929: 9-19. doi: 10.1007/978-1-62703-050-2_2
|
| [154] | Caldwell JC, Evans MV, Krishnan K (2012) Cutting Edge PBPK Models and Analyses: Providing the Basis for Future Modeling Efforts and Bridges to Emerging Toxicology Paradigms. J Toxicol 2012: 852384. |
| [155] |
Barrett JS, Della Casa Alberighi O, Laer S, et al. (2012) Physiologically based pharmacokinetic (PBPK) modeling in children. Clin Pharmacol Ther 92: 40-49. doi: 10.1038/clpt.2012.64
|
| [156] |
Bjorkman S (2005) Prediction of drug disposition in infants and children by means of physiologically based pharmacokinetic (PBPK) modelling: theophylline and midazolam as model drugs. Br J Clin Pharmacol 59: 691-704. doi: 10.1111/j.1365-2125.2004.02225.x
|
| [157] |
Vinks AA (2013) The future of physiologically based pharmacokinetic modeling to predict drug exposure in pregnant women. CPT Pharmacometrics Syst Pharmacol 2: e33. doi: 10.1038/psp.2013.9
|
| [158] |
Ruiz P, Ray M, Fisher J, et al. (2011) Development of a human Physiologically Based Pharmacokinetic (PBPK) Toolkit for environmental pollutants. Int J Mol Sci 12: 7469-7480. doi: 10.3390/ijms12117469
|
| [159] |
Hartung T, van Vliet E, Jaworska J, et al. (2012) Systems toxicology. ALTEX 29: 119-128. doi: 10.14573/altex.2012.2.119
|
| [160] |
Kitano H (2002) Computational systems biology. Nature 420: 206-210. doi: 10.1038/nature01254
|
| [161] |
Mc Auley MT, Wilkinson DJ, Jones JJ, et al. (2012) A whole-body mathematical model of cholesterol metabolism and its age-associated dysregulation. BMC Syst Biol 6: 130. doi: 10.1186/1752-0509-6-130
|
| [162] | Mc Auley MT, Proctor CJ, Corfe BM, et al. (2013) Nutrition Research and the Impact of Computational Systems Biology. l Comput Sci Syst Biol 6: 271-285. |
| [163] |
Wittig U, Rey M, Kania R, et al. (2014) Challenges for an enzymatic reaction kinetics database. FEBS J 281: 572-582. doi: 10.1111/febs.12562
|
| [164] | Gutenkunst RN, Waterfall JJ, Casey FP, et al. (2007) Universally sloppy parameter sensitivities in systems biology models. PLoS Comput Biol 3: 1871-1878. |
| [165] | Carbo A, Olivares-Villagomez D, Hontecillas R, et al. (2014) Systems modeling of the role of interleukin-21 in the maintenance of effector CD4+ T cell responses during chronic Helicobacter pylori infection. MBio 5: e01243-01214. |
| [166] |
Reibman J, Marmor M, Filner J, et al. (2008) Asthma is inversely associated with Helicobacter pylori status in an urban population. PLoS One 3: e4060. doi: 10.1371/journal.pone.0004060
|
| [167] |
Pacifico L, Osborn JF, Tromba V, et al. (2014) Helicobacter pylori infection and extragastric disorders in children: a critical update. World J Gastroenterol 20: 1379-1401. doi: 10.3748/wjg.v20.i6.1379
|
| [168] |
Wilkinson DJ (2009) Stochastic modelling for quantitative description of heterogeneous biological systems. Nat Rev Genet 10: 122-133. doi: 10.1038/nrg2509
|
| [169] |
Jaworska J, Gabbert S, Aldenberg T (2010) Towards optimization of chemical testing under REACH: a Bayesian network approach to Integrated Testing Strategies. Regul Toxicol Pharmacol 57: 157-167. doi: 10.1016/j.yrtph.2010.02.003
|
| [170] |
Chaouiya C (2007) Petri net modelling of biological networks. Brief Bioinform 8: 210-219. doi: 10.1093/bib/bbm029
|
| [171] | Hucka M, Finney A, Bornstein BJ, et al. (2004) Evolving a lingua franca and associated software infrastructure for computational systems biology: the Systems Biology Markup Language (SBML) project. Syst Biol (Stevenage) 1: 41-53. |
| [172] |
Ankley GT, Bennett RS, Erickson RJ, et al. (2010) Adverse outcome pathways: a conceptual framework to support ecotoxicology research and risk assessment. Environ Toxicol Chem 29: 730-741. doi: 10.1002/etc.34
|
| [173] |
Kimber I, Dearman RJ, Basketter DA, et al. (2014) Chemical respiratory allergy: reverse engineering an adverse outcome pathway. Toxicology 318: 32-39. doi: 10.1016/j.tox.2014.02.001
|
| [174] |
Vinken M (2013) The adverse outcome pathway concept: a pragmatic tool in toxicology. Toxicology 312: 158-165. doi: 10.1016/j.tox.2013.08.011
|
| [175] | National Research Council (2012) Exposure Science in the 21st Century: A Vision and a Strategy. Washington, DC: The National Academies Press. 196 p. |
| [176] |
Brunekreef B (2013) Exposure science, the exposome, and public health. Environ Mol Mutagen 54: 596-598. doi: 10.1002/em.21767
|
| [177] |
Bornehag CG, Blomquist G, Gyntelberg F, et al. (2001) Dampness in Buildings and Health. Indoor Air 11: 72-86. doi: 10.1034/j.1600-0668.2001.110202.x
|
| [178] | Bornehag CG, Sundell J, Bonini S, et al. (2004) Dampness in buildings as a risk factor for health effects, (EUROEXPO). A multidisciplinary review of the literature (1998-2000) on dampness and mite exposure in buildings and health effects. Indoor Air 14: 243-257. |
| [179] | Bornehag CG, Sundell J, Hägerhed-Engman L, et al. (2005) Association between ventilation rates in 390 Swedish homes and allergic symptoms in children. 15: 275-280. |
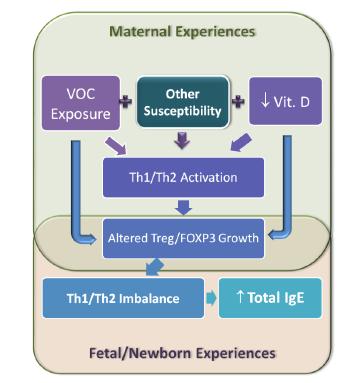
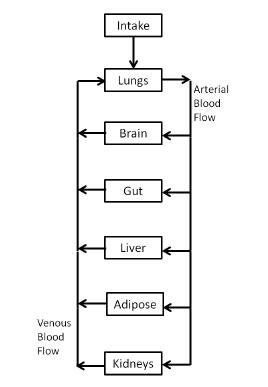
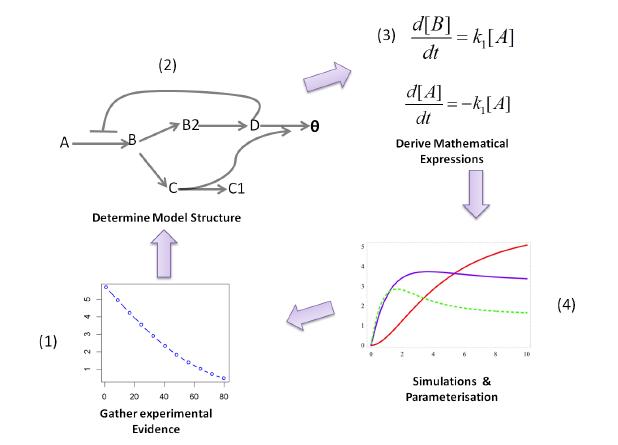
Figures(3)

Hyunok Choi, Mark T. McAuley, David A. Lawrence. Prenatal exposures and exposomics of asthma[J]. AIMS Environmental Science, 2015, 2(1): 87-109. doi: 10.3934/environsci.2015.1.87







 DownLoad:
DownLoad: