Citation: Kayla L. Penta, Diego Altomare, Devon L. Shirley, Jennifer F. Nyland. Female immune system is protected from effects of prenatal exposure to mercury[J]. AIMS Environmental Science, 2015, 2(3): 448-463. doi: 10.3934/environsci.2015.3.448

| [1] | NRC (2000) Toxicological effects of methyl mercury. Washington, DC: National Academy Press. |
| [2] |
Clarkson TW, Magos L, Myers GJ (2003) The toxicology of mercury--current exposures and clinical manifestations. N Engl J Med 349: 1731-1737. doi: 10.1056/NEJMra022471

|
| [3] |
Mergler D, Anderson HA, Chan LH, et al. (2007) Methylmercury exposure and health effects in humans: a worldwide concern. Ambio 36: 3-11. doi: 10.1579/0044-7447(2007)36[3:MEAHEI]2.0.CO;2
|
| [4] | Langworth S, Elinder CG, Sundquist KG, et al. (1992) Renal and immunological effects of occupational exposure to inorganic mercury. Br J Ind Med 49: 394-401. |
| [5] |
Grandjean P, White RF, Nielsen A, et al. (1999) Methylmercury neurotoxicity in Amazonian children downstream from gold mining. Environ Health Perspect 107: 587-591. doi: 10.1289/ehp.99107587
|
| [6] |
Grandjean P, Weihe P, White RF, et al. (1998) Cognitive performance of children prenatally exposed to "safe" levels of methylmercury. Environ Res 77: 165-172. doi: 10.1006/enrs.1997.3804
|
| [7] | Myers GJ, Davidson PW, Cox C, et al. (1995) Neurodevelopmental outcomes of Seychellois children sixty-six months after in utero exposure to methylmercury from a maternal fish diet: pilot study. Neurotoxicology 16: 639-652. |
| [8] |
Zahir F, Rizwi SJ, Haq SK, et al. (2005) Low dose mercury toxicity and human health. Environ Toxicol Pharmacol 20: 351-360. doi: 10.1016/j.etap.2005.03.007
|
| [9] | Mahaffey KR, Clickner RP, Bodurow CC (2004) Blood organic mercury and dietary mercury intake: national health and nutrition examination survey, 1999 and 2000. Environ Health Perspect 112: 562-570. |
| [10] |
Buchanan S, Anglen J, Turyk M (2015) Methyl mercury exposure in populations at risk: Analysis of NHANES 2011-2012. Environ Res 140: 56-64. doi: 10.1016/j.envres.2015.03.005
|
| [11] |
Gardner RM, Nyland JF, Evans SL, et al. (2009) Mercury induces an unopposed inflammatory response in human peripheral blood mononuclear cells in vitro. Environ Health Perspect 117: 1932-1938. doi: 10.1289/ehp.0900855
|
| [12] |
Gardner RM, Nyland JF, Silbergeld EK (2010) Differential immunotoxic effects of inorganic and organic mercury species in vitro. Toxicology Letters 198: 182-190. doi: 10.1016/j.toxlet.2010.06.015
|
| [13] |
Gardner RM, Nyland JF, Silva IA, et al. (2010) Mercury exposure, serum antinuclear/antinucleolar antibodies, and serum cytokine levels in mining populations in Amazonian Brazil: a cross-sectional study. Environ Res 110: 345-354. doi: 10.1016/j.envres.2010.02.001
|
| [14] |
Nyland JF, Fillion M, Barbosa Jr F, et al. (2011) Biomarkers of methyl mercury exposure immunotoxicity among fish consumers in Amazonian Brazil. Environ Health Perspect 119: 1733-1738. doi: 10.1289/ehp.1103741
|
| [15] |
Nyland JF, Fairweather D, Shirley DL, et al. (2012) Low-dose inorganic mercury increases severity and frequency of chronic coxsackievirus-induced autoimmune myocarditis in mice. Toxicol Sci 125: 134-143. doi: 10.1093/toxsci/kfr264
|
| [16] | Penta KL, Fairweather D, Shirley DL, et al. (2014) Low dose inorganic mercury heightens early innate response to coxsackievirus infection in female mice. Inflammation Res 64: 31-40. |
| [17] |
Harris HH, Pickering IJ, George GN (2003) The chemical form of mercury in fish. Science 301: 1203. doi: 10.1126/science.1085941
|
| [18] |
Silva IA, El Nabawi M, Hoover D, et al. (2005) Prenatal HgCl2 exposure in BALB/c mice: gender-specific effects on the ontogeny of the immune system. Dev Comp Immunol 29: 171-183. doi: 10.1016/j.dci.2004.05.008
|
| [19] |
Toomey CB, Cauvi DM, Hamel JC, et al. (2014) Cathepsin B regulates the appearance and severity of mercury-induced inflammation and autoimmunity. Toxicol Sci 142: 339-349. doi: 10.1093/toxsci/kfu189
|
| [20] |
Zhang Y, Bolivar VJ, Lawrence DA (2013) Maternal exposure to mercury chloride during pregnancy and lactation affects the immunity and social behavior of offspring. Toxicol Sci 133: 101-111. doi: 10.1093/toxsci/kft023
|
| [21] |
Ekstrand J, Nielsen JB, Havarinasab S, et al. (2010) Mercury toxicokinetics--dependency on strain and gender. Toxicol Appl Pharmacol 243: 283-291. doi: 10.1016/j.taap.2009.08.026
|
| [22] |
Clarkson TW (1997) The toxicology of mercury. Crit Rev Clin Lab Sci 34: 369-403. doi: 10.3109/10408369708998098
|
| [23] |
Thuvander A, Sundberg J, Oskarsson A (1996) Immunomodulating effects after perinatal exposure to methylmercury in mice. Toxicology 114: 163-175. doi: 10.1016/S0300-483X(96)03486-5
|
| [24] |
Virtanen JK, Rissanen TH, Voutilainen S, et al. (2007) Mercury as a risk factor for cardiovascular diseases. Journal of Nutritional Biochemistry 18: 75-85. doi: 10.1016/j.jnutbio.2006.05.001
|
| [25] | Nielsen JB, Hultman P (2002) Mercury-induced autoimmunity in mice. Environ Health Perspect 110 Suppl 5: 877-881. |
| [26] |
Pollard KM, Pearson DL, Hultman P, et al. (2001) Xenobiotic acceleration of idiopathic systemic autoimmunity in lupus- prone bxsb mice. Environ Health Perspect 109: 27-33. doi: 10.1289/ehp.01109s127
|
| [27] | Martinez FO, Gordon S (2014) The M1 and M2 paradigm of macrophage activation: time for reassessment. F1000Prime Rep 6: 13. |
| [28] |
Mantovani A, Sica A, Sozzani S, et al. (2004) The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol 25: 677-686. doi: 10.1016/j.it.2004.09.015
|
| [29] |
Chattopadhyay K, Ramagopal UA, Brenowitz M, et al. (2008) Evolution of GITRL immune function: murine GITRL exhibits unique structural and biochemical properties within the TNF superfamily. Proc Natl Acad Sci U S A 105: 635-640. doi: 10.1073/pnas.0710529105
|
| [30] |
Zhou Z, Tone Y, Song X, et al. (2008) Structural basis for ligand-mediated mouse GITR activation. Proc Natl Acad Sci U S A 105: 641-645. doi: 10.1073/pnas.0711206105
|
| [31] |
Nocentini G, Ronchetti S, Cuzzocrea S, et al. (2007) GITR/GITRL: more than an effector T cell co-stimulatory system. Eur J Immunol 37: 1165-1169. doi: 10.1002/eji.200636933
|
| [32] | Leavesley DI, Kashyap AS, Croll T, et al. (2013) Vitronectin--master controller or micromanager? IUBMB Life 65: 807-818. |
| [33] | Seiffert D, Geisterfer M, Gauldie J, et al. (1995) IL-6 stimulates vitronectin gene expression in vivo. J Immunol 155: 3180-3185. |
| [34] |
Villanueva MBG, Koizumi S, Jonai H (2000) Cytokine production by human peripheral blood mononuclear cells after exposure to heavy metals. Journal of Health Science 46: 358-362. doi: 10.1248/jhs.46.358
|
| [35] |
Preissner KT (1991) Structure and biological role of vitronectin. Annu Rev Cell Biol 7: 275-310. doi: 10.1146/annurev.cb.07.110191.001423
|
| [36] |
Hezova R, Slaby O, Faltejskova P, et al. (2010) microRNA-342, microRNA-191 and microRNA-510 are differentially expressed in T regulatory cells of type 1 diabetic patients. Cell Immunol 260: 70-74. doi: 10.1016/j.cellimm.2009.10.012
|
| [37] |
Moore KW, de Waal Malefyt R, Coffman RL, et al. (2001) Interleukin-10 and the interleukin-10 receptor. Annu Rev Immunol 19: 683-765. doi: 10.1146/annurev.immunol.19.1.683
|
| [38] |
Saraiva M, O'Garra A (2010) The regulation of IL-10 production by immune cells. Nat Rev Immunol 10: 170-181. doi: 10.1038/nri2711
|
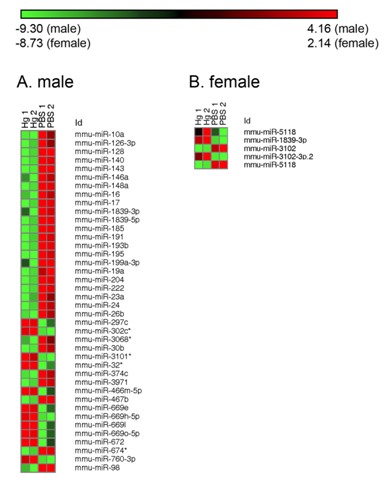

Figures(6) / Tables(2)

Kayla L. Penta, Diego Altomare, Devon L. Shirley, Jennifer F. Nyland. Female immune system is protected from effects of prenatal exposure to mercury[J]. AIMS Environmental Science, 2015, 2(3): 448-463. doi: 10.3934/environsci.2015.3.448







 DownLoad:
DownLoad: