Citation: Xiu Zhang, Nhuan P. Nghiem. Pretreatment and Fractionation of Wheat Straw for Production of Fuel Ethanol and Value-added Co-products in a Biorefinery[J]. AIMS Bioengineering, 2014, 1(1): 40-52. doi: 10.3934/bioeng.2014.1.40

| [1] | FAOSTAT, Food and Agriculture Organization of the United Nations, 2014. Available from: http://faostat3.fao.org/faostat-gateway/go/to/download/Q/QC/E. |
| [2] |
Kim S, Dale BE (2004) Global potential bioethanol production from wasted crops and crop residues. Biomass Bioenerg 26: 361-375. doi: 10.1016/j.biombioe.2003.08.002

|
| [3] | Padgitt M, Newton D, Penn R, et al. (2013). Production Practices for Major Crops in U.S. Agriculture, 1990-97, USDA Economic Research Service. Available from: http://www.ers.usda.gov/publications/sb-statistical-bulletin/sb969.aspx#.U7rDCrH5eYc. |
| [4] | Drapcho CM, Nghiem NP, Walker TH (2008) Biofuels Engineering Process Technology, New York: McGraw-Hill, 69-103. |
| [5] |
Saha BC, Iten LB, Cotta MA, et al. (2005) Dilute acid pretreatment, enzymatic saccharification and fermentation of wheat straw to ethanol. Process Biochem 40: 3693-3700. doi: 10.1016/j.procbio.2005.04.006
|
| [6] |
Saha BC, Cotta MA (2006) Ethanol production from alkaline peroxide pretreated enzymatically saccharified wheat straw. Biotechnol Prog 22: 449-453. doi: 10.1021/bp050310r
|
| [7] |
Ballesteros I, Negro MJ, Oliva JM, et al. (2006) Ethanol production from steam-exploded wheat straw. Appl Biochem Biotechnol 130: 496-508. doi: 10.1385/ABAB:130:1:496
|
| [8] |
Hongzhang C, Liying L (2007) Unpolluted fractionation of wheat straw by steam explosion and ethanol extraction. Bioresource Technol 98: 666-676. doi: 10.1016/j.biortech.2006.02.029
|
| [9] |
Zhu S, Wu Y, Yu Z, et al. (2006) Production of ethanol from microwave-assisted alkali pretreated wheat straw. Process Biochem 41: 869-873. doi: 10.1016/j.procbio.2005.10.024
|
| [10] |
Klinke HB, Olsson L, Thomsen AB, et al. (2003) Potential inhibitors from wet oxidation of wheat straw and their effect on ethanol production of Saccharomyces cerevisiae: Wet oxidation and fermentation by yeast. Biotechnol Bioeng 81: 738-747. doi: 10.1002/bit.10523
|
| [11] |
Kim TH, Taylor F, Hicks KB (2008) Bioethanol production from barley hull using SAA (soaking in aqueous ammonia) pretreatment. Bioresource Technol 99: 5694-5702. doi: 10.1016/j.biortech.2007.10.055
|
| [12] |
Nghiem NP, Kim TH, Yoo GC, et al. (2013) Enzymatic fractionation of pretreated barley straw for production of fuel ethanol and astaxanthin as a value-added co-product. Appl Biochem Biotechnol 171: 341-351. doi: 10.1007/s12010-013-0374-0
|
| [13] | Nghiem NP, Kim TH, Hicks KB (2012) Method for improving the bioavailability of polysaccharides in lignocellulosic materials. US Patent 8,202,970. |
| [14] |
Kim TH, Lee YY (2005) Pretreatment of corn stover by soaking in aqueous ammonia. Appl Biochem Biotechnol 124: 1119-1132. doi: 10.1385/ABAB:124:1-3:1119
|
| [15] | Kim TH, Lee YY (2007) Pretreatment of corn stover by soaking in aqueous ammonia at moderate temperatures. Appl Biochem Biotechnol 137: 81-92. |
| [16] |
Leathers TD (2003) Bioconversions of maize residues to value-added coproducts using yeast-like fungi. FEMS Yeast Res 3: 133-140. doi: 10.1016/S1567-1356(03)00003-5
|
| [17] |
Todd Lorenz R, Cysewski GR (2000) Commercial potential for Haematococcus microalgae as a natural source of astaxanthin. Trends Biotechnol 18: 160-167. doi: 10.1016/S0167-7799(00)01433-5
|
| [18] |
Guerin M, Huntley ME, Olaizola M (2003) Haematococcus astaxanthin: applications for human health and nutrition. Trends Biotechnol 21: 210-216. doi: 10.1016/S0167-7799(03)00078-7
|
| [19] | Higuera-Ciapara I, Felix-Valenzuela L, Goycoolea FM (2007) Astaxanthin: A review of its chemistry and applications. Crit Rev Food Sci Nutrition 46: 185-196. |
| [20] |
Nghiem NP, Montanti J, Johnston DB (2009) Production of astaxanthin from corn fiber as a value-added co-product of fuel ethanol fermentation. Appl Biochem Biotechnol 154: 48-58. doi: 10.1007/s12010-008-8399-5
|
| [21] | Prakasham RS, Rao RS, Hobbs PJ (2009) Current trends in biotechnological production of xylitol and future prospects. Curr Trends Biotechnol Pharm 3: 8-36. |
| [22] | Granström TB, Izumori K, Leisola M (2007) A rare sugar xylitol. Part II: biotechnological production and future applications of xylitol. Appl Microbiol Biotechnol 74: 273-276. |
| [23] |
Rivas B, Torre P, Dominguez JM, et al. (2009) Maintenance and growth requirements in the metabolism of Debaryomyces hansenii performing xylose-to-xylitol bioconversion in corncob hemicellulose hydrolysate. Biotechnol Bioeng 102: 1062-1073. doi: 10.1002/bit.22155
|
| [24] | Sluiter A, Hames B, Ruiz R, et al. (2011) Determination of structural carbohydrates and lignin in biomass. Technical report NREL/TP-510-42618. Available from: http://www.nrel.gov/biomass/pdfs/42618.pdf |
| [25] |
Yoo GC, Nghiem NP, Hicks KB, et al. (2013) Maximum production of fermentable sugars from barley straw using optimized soaking in aqueous ammonia (SAA) pretreatment. Appl Biochem Biotechnol 169: 2430-2441. doi: 10.1007/s12010-013-0154-x
|
| [26] |
Tochampa W, Sirisansaneeyakul S, Vanichsriratana W, et al. (2005) A model of xylitol production by the yeast Candida mogii. Bioprocess Biosys Eng 28: 175-183. doi: 10.1007/s00449-005-0025-0
|
| [27] | Collura MA, Luyben WL (1988) Energy-saving distillation designs in ethanol production, Ind Eng Chem Res 27: 1686-1696. |
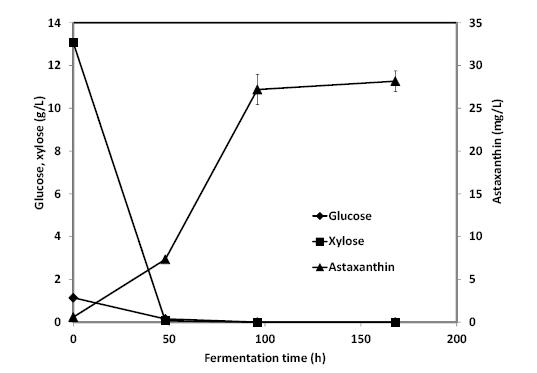
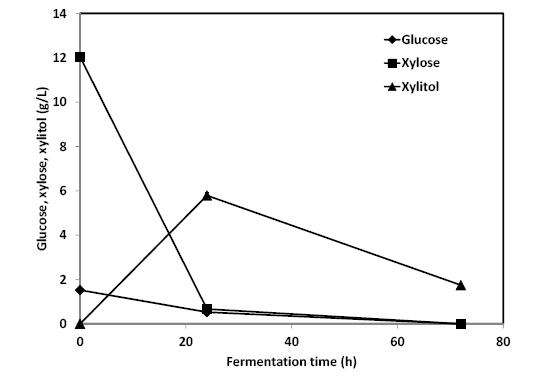
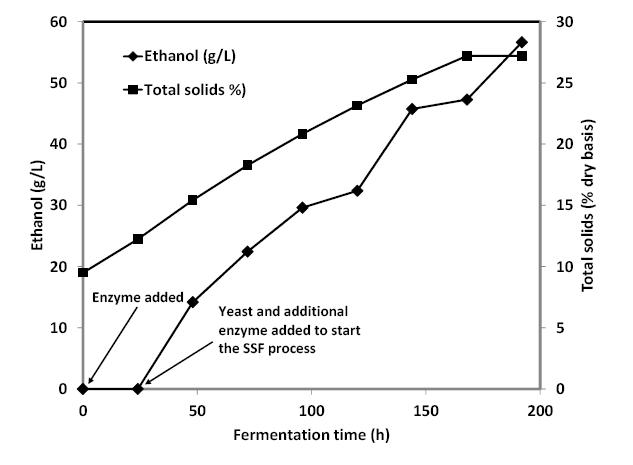
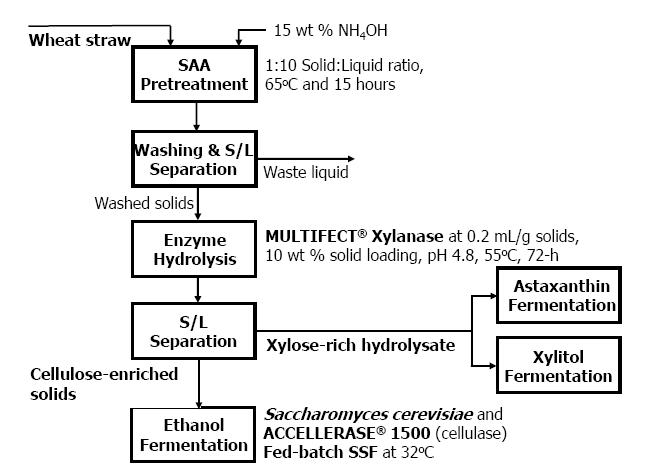
Figures(4) / Tables(1)

Xiu Zhang, Nhuan P. Nghiem. Pretreatment and Fractionation of Wheat Straw for Production of Fuel Ethanol and Value-added Co-products in a Biorefinery[J]. AIMS Bioengineering, 2014, 1(1): 40-52. doi: 10.3934/bioeng.2014.1.40







 DownLoad:
DownLoad: