Citation: Scott Rollo, Anca Gaston, Harry Prapavessis. Cognitive and Motivational Factors Associated with Sedentary Behavior: A Systematic Review[J]. AIMS Public Health, 2016, 3(4): 956-984. doi: 10.3934/publichealth.2016.4.956

| [1] |
Rezende, LF, Rodrigues Lopes M, Rey-López JP, et al. (2014) Sedentary behavior and health outcomes: An overview of systematic reviews. PLoS ONE 9: e105620. doi: 10.1371/journal.pone.0105620

|
| [2] | Sedentary Behavior Research Network (2012) Letter to the editor: standardized use of the terms "sedentary" and "sedentary behaviours". Appl Physiol Nutr Metab 37: 540-542. |
| [3] | Colley RC, Garriguet D, Janssen I, et al. (2011) Physical activity of Canadian adults: accelerometer results from the 2007 to 2009 Canadian health measures survey. Health Rep 22: 7-14. |
| [4] | Matthews CE, Chen KY, Freedson PS, et al. (2008) Amount of time spent in sedentary behaviors in the United States, 2003–2004. Am J Epidemiol 167: 875-881. |
| [5] | Owen N, Sugiyama T, Eakin EE, et al. (2011) Adults' sedentary behavior: determinants and interventions. Am J Prev Med 41: 189-196. |
| [6] | Sallis JF, Owen N, Fisher EB (2008) Ecological models of health behaviour, In: Glanz K, Rimer BK, & Viswanath K, Health behavior and health education: Theory, research, and practice, 4 Eds., San Francisco, CA: Jossey-Bass, 465-486. |
| [7] | Forgas JP (2008) Affect and cognition. Perspect Psychol Sci 3: 94-101. |
| [8] | Hilgard ER (1980) The trilogy of mind: Cognition, affection, and conation. J Hist Behav Sci 16: 107-117. |
| [9] | Rosenstock I (1974) The health belief model and preventive health behavior. Health Educ Monogr 2: 355-385. |
| [10] | Ajzen I & Fishbein M (1977) Attitude-behavior relations: A theoretical analysis and review of empirical research. Psychol Bull 84: 888. |
| [11] | Ajzen I (1991) The theory of planned behavior. Organ Behav Hum Decis Process 50: 179-211. |
| [12] | Rogers RW (1975) A protection motivation theory of fear appeals and attitude change. J Psychol 91: 93-114. |
| [13] | Bandura A (1977) Self-efficacy: toward a unifying theory of behavioral change. Psychol Rev 84: 191. |
| [14] | Schwarzer R (2008) Modeling health behavior change: how to predict and modify the adoption and maintenance of health behaviors. Appl Psychol 57: 1-29. |
| [15] | Deci EL & Ryan RM (2002) Handbook of self-determination research. Rochester, NY: University of Rochester Press. |
| [16] | Armitage CJ & Conner M (2000) Social cognition models and health behavior: A structured review. Psychol Health 15: 173-189. |
| [17] | Rhodes RE, Mark RS, Temmel CP (2012) Adult sedentary behavior: A systematic review. American J Prev Med 42: e3-e28. |
| [18] | Owen N, Salmon J, Koohsari MJ, et al. (2014) Sedentary behaviour and health: mapping environmental and social contexts to underpin chronic disease prevention. Br J Sports Med 48: 174-177. |
| [19] | Salmon J, Tremblay MS, Marshall SJ, et al. (2011) Health risks, correlates, and interventions to reduce sedentary behavior in young people. Am J Prev Med 41: 197-206. |
| [20] | Uijtdewilligen L, Twisk JWR, van der Horst K, et al. (2012) Determinants of physical activity and sedentary behaviour in young people: a review and quality synthesis of prospective studies. Br J Sports Med 45: 896-905. |
| [21] | Atkin AJ, Corder K, Goodyer I, et al. (2015) Perceived family functioning and friendship quality: cross-sectional association with physical activity and sedentary behaviours. Int J Behav Nutr Phys Act 12: 23. |
| [22] | Bai Y, Chen S, Vazou S, et al. (2015) Mediated effects of perceived competence on youth physical activity and sedentary behaviour. Res Q Exerc Sport 86: 406-413. |
| [23] | Busschaert C, De Bourdeaudhuij I, Van Cauwenberg J, et al. (2016) Intrapersonal, social-cognitive and physical environmental variables related to context-specific sitting time in adults: a one-year follow-up study. Int J Behav Nutr Phys Act 13: 28. |
| [24] | Chang AK & Sok SR (2015) Predictors of sedentary behavior in elderly Koreans with hypertension. J Nurs Res 23: 262-270. |
| [25] | Conroy DE, Maher JP, Elavsky S, et al. (2013) Sedentary behaviour as a daily process regulated by habits and intentions. Health Psychol 32: 1149-1157. |
| [26] | De Cocker K, Duncan MJ, Short C, et al. (2014) Understanding occupational sitting: Prevalence, correlates and moderating effects in Australian employees. Prev Med 67: 288-294. |
| [27] | Gaston A, De Jesus S, Markland D, et al. (2016) I sit because I have fun when I do so! Using self-determination theory to understand sedentary behavior motivation. Health Psychol Behav Med (In Press). |
| [28] | Gebremariam MK, Totland TH, Andersen LF, et al. (2012) Stability and change in screen-based sedentary behaviours and associated factors among Norwegian children in the transition between childhood and adolescence. BMC Public Health 12: 104. |
| [29] | Ham OK, Sung KM, Kim HK (2013) Factors associated with screen time among school-age children in Korea. J Sch Nurs 29: 425-434. |
| [30] | He M, Piché L, Beynon C, et al. (2010) Screen-related sedentary behaviors: Children’s and parents’ attitudes, motivations, and practices. J Nutr Educ Behav 42: 17-25. |
| [31] | Hoyos Cillero I, Jago R, Sebire S (2011) Individual and social predictors of screen-viewing among Spanish school children. Eur J Pediatr 170: 93-102. |
| [32] | Huang WY, Wong SH, Salmon J (2013) Correlates of physical activity and screen-based behaviors in Chinese children. J Sci Med Sport 16: 509-514. |
| [33] | Janssen X, Basterfield L, Parkinson KN, et al. (2015) Determinants of changes in sedentary time and breaks in sedentary time among 9 and 12 years old children. Prev Med Rep 2: 880-885. |
| [34] | Kremers SPJ & Brug J (2008) Habit strength of physical activity and sedentary behavior among children and adolescents. Pediatr Exerc Sci 20: 5-17. |
| [35] | Lowe SS, Danielson B, Beaumont C, et al. (2015) Correlates of objectively measured sedentary behavior in cancer patients with brain metastases: an application of the theory of planned behavior. Psycho-Oncology 24: 757-762. |
| [36] | Maher JP & Conroy DE (2015) Habit strength moderates the effects of daily action planning prompts on physical activity but not sedentary behavior. J Sport Exerc Psychol 37: 97-107. |
| [37] | Maher JP & Conroy DE (2016) A dual-process model of older adults’ sedentary behavior. Health Psychol 35: 262-272. |
| [38] | Norman GJ, Schmid BA, Sallis JF, et al. (2005) Psychosocial and environmental correlates of adolescent sedentary behaviors. Pediatrics 116: 908-916. |
| [39] | Prapavessis H, Gaston A, DeJesus S (2015) The theory of planned behavior as a model for understanding sedentary behavior. Psychol Sport Exerc 19: 23-32. |
| [40] | Quartiroli A & Maeda H (2014) Self-determined engagement in physical activity and sedentary behaviors of US college students. Int J Exerc Sci 7: 87-97. |
| [41] | Rhodes RE & Dean RN (2009) Understanding physical inactivity: Prediction of four sedentary leisure behaviors. Leis Sci 31: 124-135. |
| [42] | Salmon J, Owen N, Crawford D, et al. (2003) Physical activity and sedentary behavior: A population-based study of barriers, enjoyment, and preference. Health Psychol 22: 178-188. |
| [43] | Van Dyck D, Cardon G, Deforche B, et al. (2011) Socio-demographic, psychosocial and home-environmental attributes associated with adults’ domestic screen time. BMC Public Health 11: 668. |
| [44] | Wallmann-Sperlich B, Bucksch J, Schneider S, et al. (2014) Socio-demographic, behavioural and cognitive correlates of work-related sitting time in German men and women. BMC Public Health 14: 1259. |
| [45] | Wong TS, Gaston A, DeJesus S, et al. (2016) The utility of a protection motivation theory framework for understanding sedentary behaviour. Health Psychol Behav Med 4: 29-48. |
| [46] | Rhodes RE & Nigg CR (2011) Advancing physical activity theory: A review and future directions. Exerc Sport Sci Rev 39: 113-119. |
| [47] | Ajzen I & Fishbein M (2005) The influence of attitudes on behavior. In: Albarracín D, Johnson BT, & Zanna MP, The Handbook of Attitudes, Mahwah, NJ: Erlbaum, 173-221. |
| [48] | Manning M (2009) The effects of subjective norms on behavior in the theory of planned behavior: a meta-analysis. Br J Soc Psychol 48: 649e705. |
| [49] | Aarts H, Paulussen T, Schaalma H (1997) Physical exercise habit: On the conceptualization and formation of habitual health behaviours. Health Educ Res 12: 363-374. |
| [50] | Grove JR & Zillich I (2003) Conceptualisation and measurement of habitual exercise. In: Katsikitis M., Proceedings of the 38th Annual Conference of the Australian Psychological Society, Melbourne: Australian Psychological Society, 88-92. |
| [51] | Mullan E, Markland D, Ingledew DK (1997) A graded conceptualisation of self- determination in the regulation of exercise behavior: Development of a measure using confirmatory factor analytic procedures. Personal Individ Differ 23: 745-752. |
| [52] | Carr LJ, Karvinen K, Peavler M, et al. (2013) Multicomponent intervention to reduce daily sedentary time: a randomized controlled trial. BMJ Open Access 3: e003261. |
| [53] | Gardiner PA, Eakin EG, Healy GN, et al. (2011) Feasibility of reducing older adults’ sedentary time. Am J Prev Med 41: 174-177. |
| [54] | Gaston A & Prapavessis H (2014) Using a combine protection motivation theory and health action process approach intervention to promote exercise during pregnancy. J Behav Med 37: 173-184. |
| [55] | Deliens T, Deforche B, De Bourdeaudhuij I, et al. (2015) Determinants of physical activity and sedentary behavior in university students: a qualitative study using focus group discussion. BMC Public Health 15: 201. |
| [56] | Uijtdewilligen L, Singh AS, Chinapaw MJM, et al. (2015) Person-related determinants of TV viewing and computer time in a cohort of young Dutch adults: Who sits the most? Scand J Med Sci Sports 25: 716-723. |
| [57] | Breland JY, Fox AM, Horowitz CR (2013) Screen time, physical activity and depression risk in minority women. Ment Health Phys Act 6: 10-15. |
| [58] | Hagger MS, Chatzisarantis NLD, Biddle SJH (2002) A meta-analytic review of the theories of reasoned action and planned behavior in physical activity: predictive validity and the contribution of additional variables. J Sport Exerc Psychol 24: 3e32. |
| [59] | Moher D, Liberati A, Tetzlaff J, et al. (2010) Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. Int J Surg 8: 336-341. |
| [60] | Downs SH & Black N (1998) The feasibility of creating a checklist for the assessment of the methodological quality both of randomised and non-randomised studies of health care interventions. J Epidemiol Community Health 52: 377-384. |
| [61] | Brocklebank LA, Falconer CL, Page AS, et al. (2015) Accelerometer-measured sedentary time and cardiometabolic biomarkers: A systematic review. Prev Med 76: 92-102. |
| [62] | Healy GN, Matthews CE, Dunstan DW, et al. (2011) Sedentary time and cardio-metabolic biomarkers in US adults: NHANES 2003-06. Eur Heart J 32: 590-597. |
| [63] |
Mansoubi M, Pearson N, Biddle S, et al. (2014) The relationship between sedentary behaviour and physical activity in adults: A systematic review. Prev Med 69: 28-35. doi: 10.1016/j.ypmed.2014.08.028
|
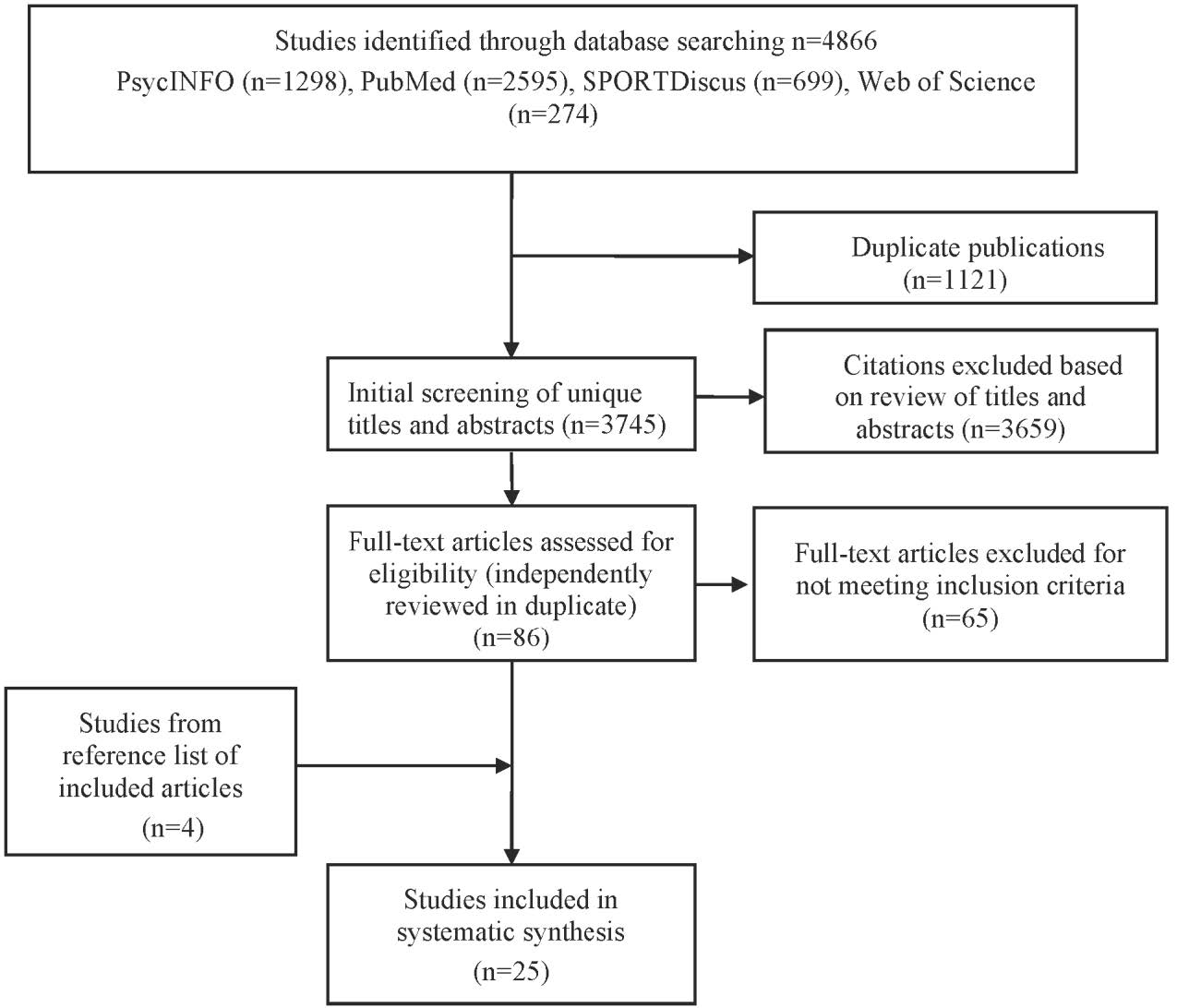

Figures(2) / Tables(2)

Scott Rollo, Anca Gaston, Harry Prapavessis. Cognitive and Motivational Factors Associated with Sedentary Behavior: A Systematic Review[J]. AIMS Public Health, 2016, 3(4): 956-984. doi: 10.3934/publichealth.2016.4.956







 DownLoad:
DownLoad: