Citation: Mijeong Kim, Seungtaek Jeong, Jong-min Yeom, Hyun-ok Kim, Jonghan Ko. Determination of rice canopy growth based on high resolution satellite images: a case study using RapidEye imagery in Korea[J]. AIMS Environmental Science, 2016, 3(4): 631-645. doi: 10.3934/environsci.2016.4.631

| [1] | Haboudane D, Miller JR, Tremblay N, et al. (2002) Integrated narrow-band vegetation indices for prediction of crop chlorophyll content for application to precision agriculture. Remote sens environ 81: 416-426. |
| [2] | Berni J, Zarco-Tejada PJ, Suárez L, et al. (2009) Thermal and narrowband multispectral remote sensing for vegetation monitoring from an unmanned aerial vehicle. IEEE Trans Geosci Remote Sens 47: 722-738. |
| [3] | Bauer ME, Cipra JE (1973) Identification of Agricultural Crops by Computer Processing of ERTS MSS Data. Symp. on Significant Results Obtained from the Earth Resources Technology Satellite-1. NASA SP-327. NASA Goddard Space Flight Center: 205-212. |
| [4] |
Lawrence SB, Xuemin J, Brian G, et al. (2012) Quick atmospheric correction code: algorithm description and recent upgrades. Opt Eng 51: 111719. doi: 10.1117/1.OE.51.11.111719

|
| [5] | Richter R (2005) Hyperspectral sensors for military applications. NASA technical report RTO-MP-SET-094, 2005. |
| [6] |
Gao BC, Heidebrecht KB, Goetz AFH (1993) Derivation of scaled surface reflectance from AVIRIS data. Remote Sens Environ 44: 145-163. doi: 10.1016/0034-4257(93)90013-N
|
| [7] | Richter R (1996) A spatially adaptive fast atmosphere correction algorithm. Int J Remote Sens 11: 159-166. |
| [8] | Adler-Golden S, Berk A, Bernstein LS, et al. (1998) FLAASH, A MODTRAN4 atmospheric correction package for hyperspectral data retrievals and simulations. In Proc 7th Ann JPL Airborne Earth Science Workshop: 9-14. |
| [9] | Tuominen J, Lipping T (2011) Detection of environmental change using hyperspectral remote sensing at Olkiluoto repository site. Working Report, Posiva Oy, Eurajoki, Finland, 31-34. |
| [10] | Jones HG, Vaughan RA (2010) Remote sensing of vegetation: principles, techniques, and applications. Oxford university press. |
| [11] | Richards JA, Jia X (2006) Remote sensing digital image analysis, 4th ed. Berlin et al. Springer 78: 193. |
| [12] |
Kim H, Yeom J (2014) Sensitivity of vegetation indices to spatial degradation of RapidEye imagery for paddy rice detection: a case study of South Korea. GISci Remote Sens 52: 1-17. doi: 10.1109/TGRS.2013.2290671
|
| [13] | RapidEye AG (2011) Satellite imagery product specifications, Version 2.1. |
| [14] |
Bernstein LS, Jin X, Gregor B, et al. (2012) Quick atmospheric correction code: algorithm description and recent upgrades. Optical engineering 51: 111719-1. doi: 10.1117/1.OE.51.11.111719
|
| [15] | ENVI (2009) Atmospheric Correction Module: QUAC and FLAASH User's Guide, Version 4. 7. ITT Visual Information Solutions, Boulder, CO. |
| [16] |
Black M, Fleming A, Riley T, et al. (2014) On the atmospheric correction of antarctic airborne hyperspectral data. Remote Sensing 6: 4498-4514. doi: 10.3390/rs6054498
|
| [17] | ERDAS, Geosystems (2009) ATCOR for ERDAS IMAGINE 2010—Haze reduction, atmospheric and topographic correction—User manual ATCOR 2 and ATCOR 3. ERDAS Imagine 1–58. ERDAS–GeoSystems. |
| [18] | Liang S (2005) Quantitative remote sensing of land surfaces. John Wiley & Sons. Inc., New York. |
| [19] | Mujumdar PP, Kumar DN (2013) Floods in a changing climate: hydrologic modeling. International Hydrology Series, Cambridge University Press, Cambridge. |
| [20] | Lillesand TM, Kiefer RW, Chipman JW (2004) Remote sensing and image interpretation (No. Ed. 5). John Wiley & Sons. Inc. New York. |
| [21] |
Xiao X, Boles S, Frolking S, et al. (2006) Mapping paddy rice agriculture in South and Southeast Asia using multi-temporal MODIS images. Remote Sens Environ 100: 95-113. doi: 10.1016/j.rse.2005.10.004
|
| [22] |
Nash J, Sutcliffe JV (1970) River flow forecasting through conceptual models part I—A discussion of principles. J hydrology 10: 282-290. doi: 10.1016/0022-1694(70)90255-6
|
| [23] | ENVI (2004) ENVI user’s guide. Research system Inc. Available from: http://aviris.gl.fcen.uba.ar/Curso_SR/biblio_sr/ENVI_userguid.pdf. |
| [24] | Thomas V, Treitz P, Jelinski D, et al. (2002) Image classification of a northern peatland complex using spectral and plant community data. Remote Sens Environ 84: 83-99. |
| [25] |
Moses WJ, Gitelson AA, Perk RL, et al. (2012) Estimation of chlorophyll-a concentration in turbid productive waters using airborne hyperspectral data. Water research 46: 993-1004. doi: 10.1016/j.watres.2011.11.068
|
| [26] | Jeong ST, Jang KC, Hong SY, et al. (2011) Detection of irrigation timing and the mapping of paddy cover in Korea using MODIS images data. Kor J Agric Forest Meteo 13: 69-78. |
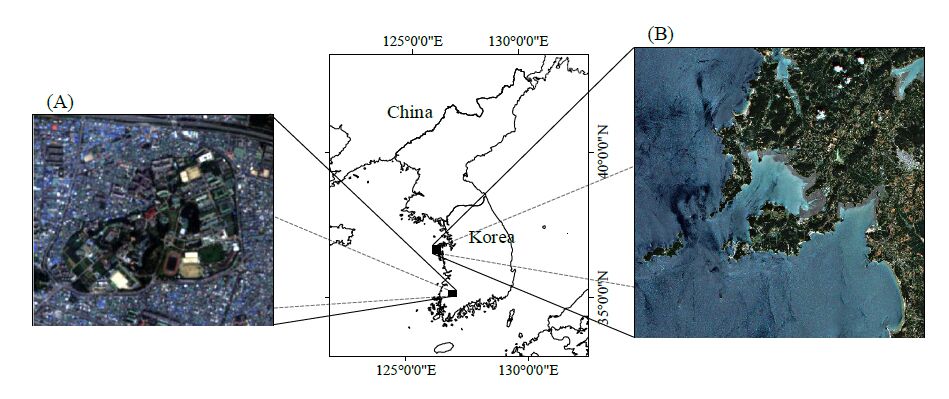
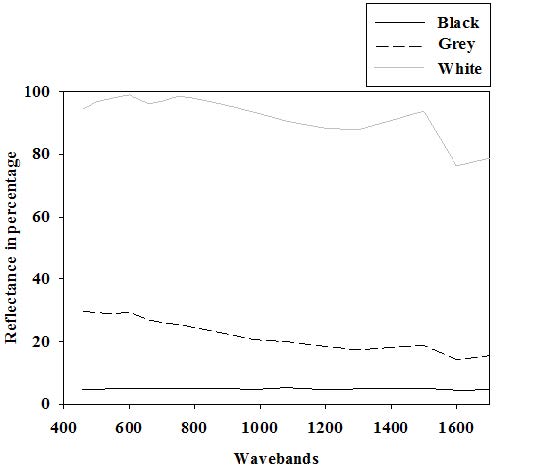
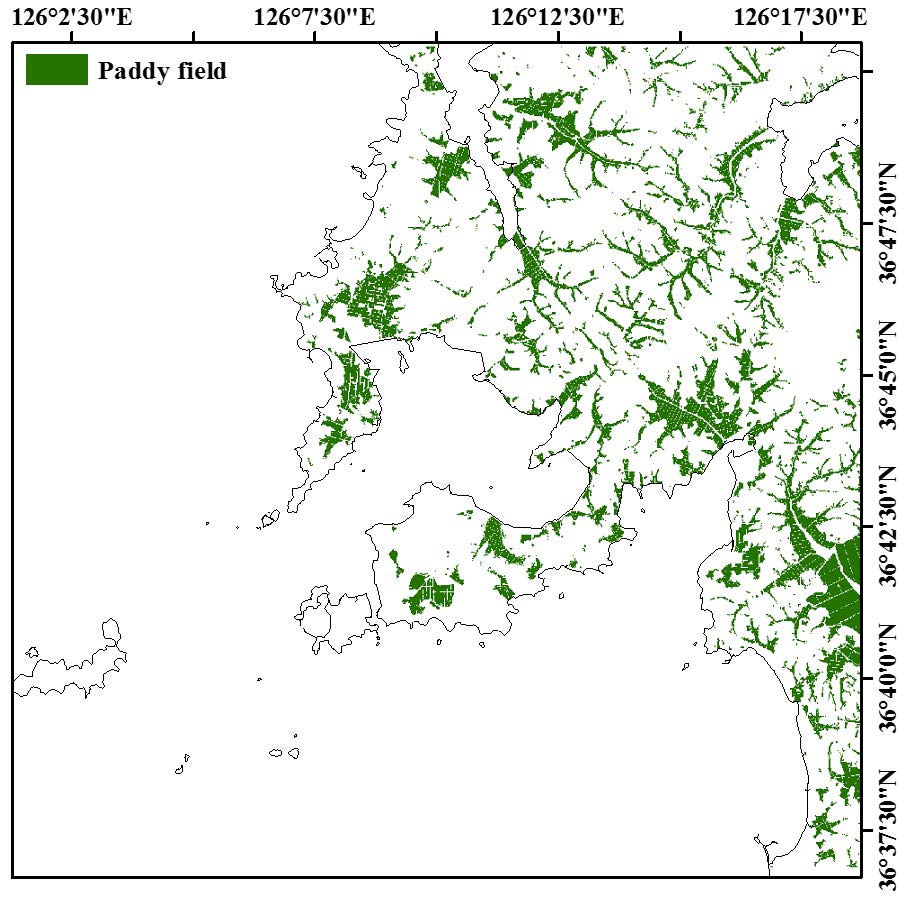
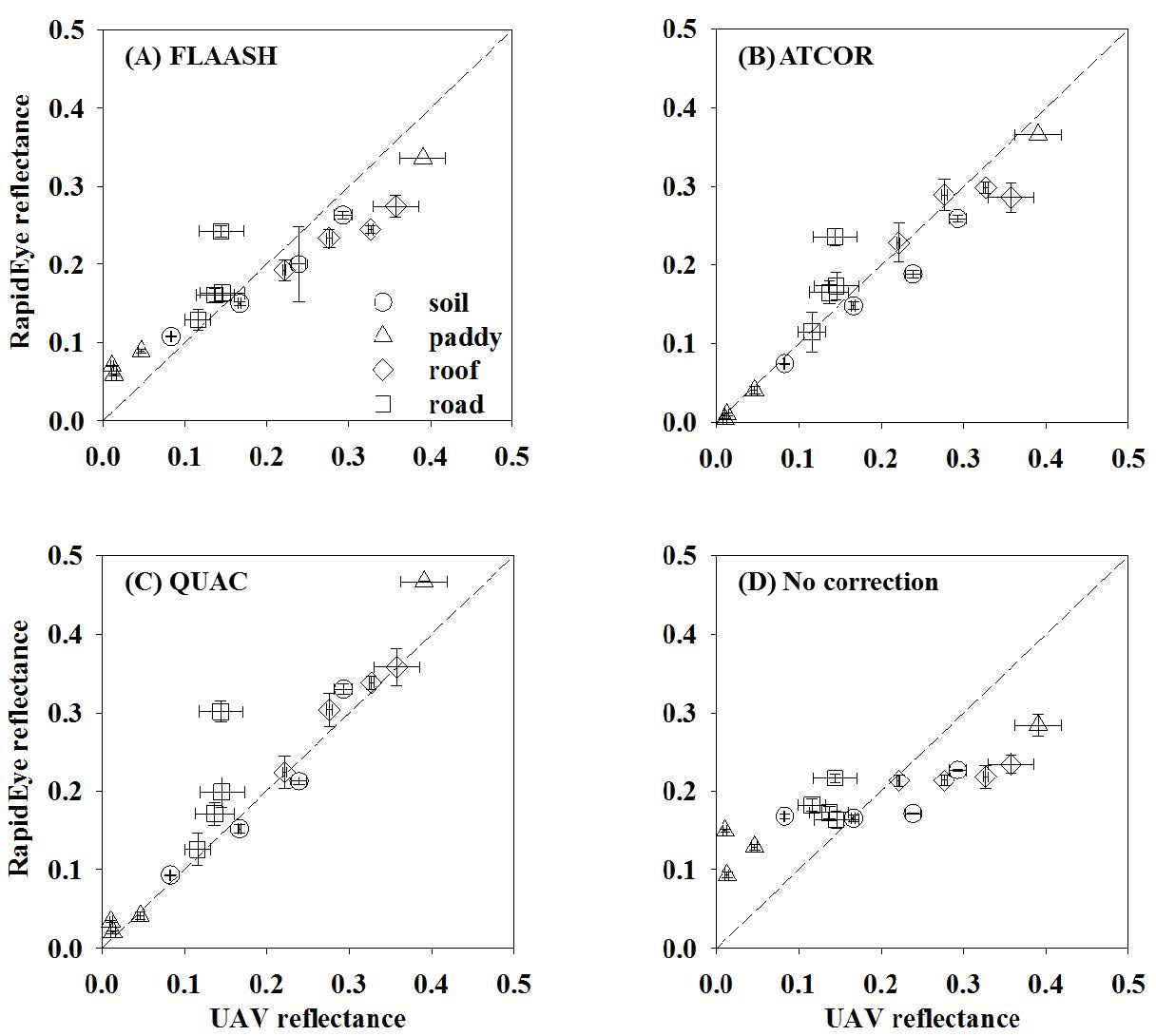
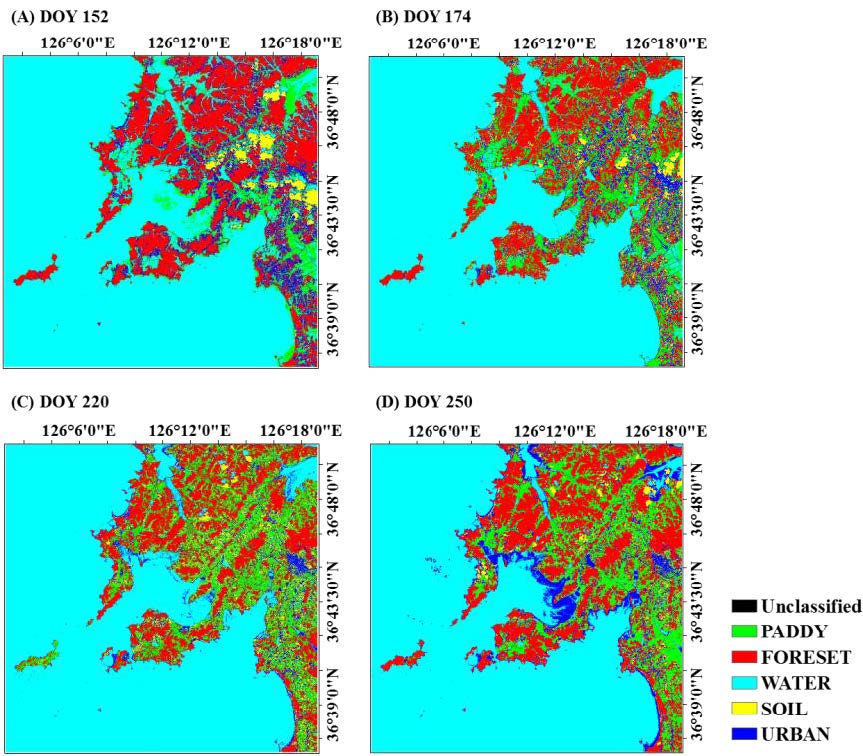
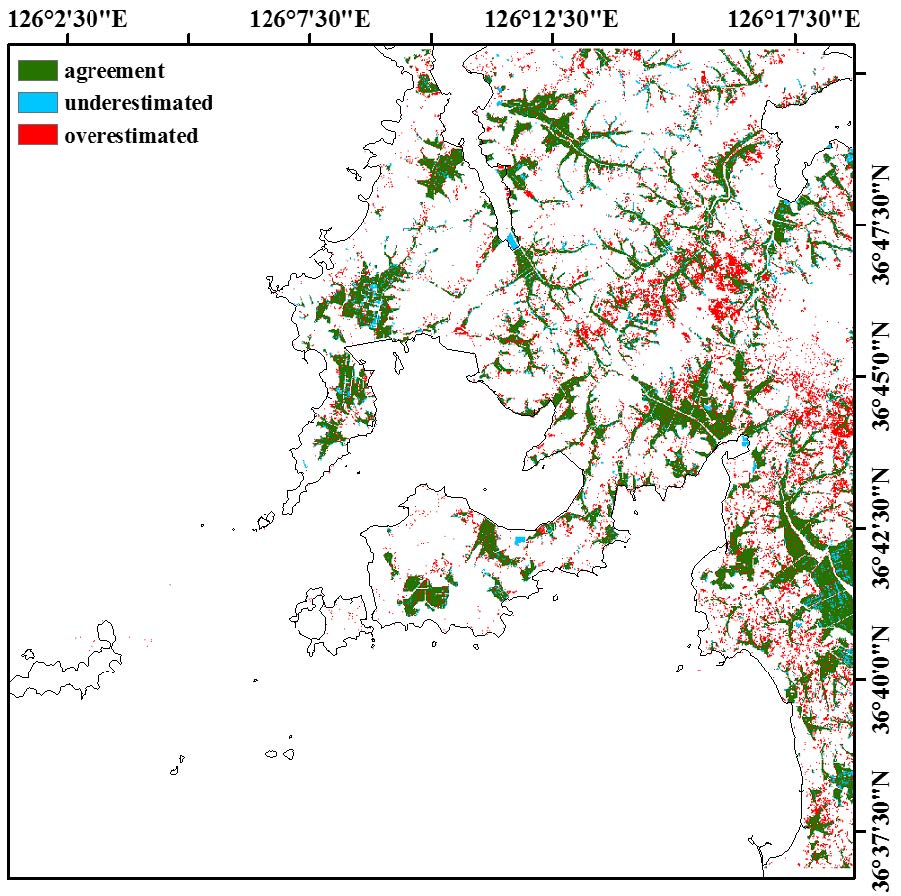
Figures(6) / Tables(6)

Mijeong Kim, Seungtaek Jeong, Jong-min Yeom, Hyun-ok Kim, Jonghan Ko. Determination of rice canopy growth based on high resolution satellite images: a case study using RapidEye imagery in Korea[J]. AIMS Environmental Science, 2016, 3(4): 631-645. doi: 10.3934/environsci.2016.4.631







 DownLoad:
DownLoad: