Citation: Teresa M. Fonovich. Phospholipid synthetic and turnover pathways elicited upon exposure to different xenobiotics[J]. AIMS Molecular Science, 2020, 7(3): 211-228. doi: 10.3934/molsci.2020010

| [1] |
Lou HY, Zhao W, Li X, et al. (2019) Membrane curvature underlies actin reorganization in response to nanoscale surface topography. Procced Natl Acad Sci 116: 23143-23151. doi: 10.1073/pnas.1910166116

|
| [2] |
Jensen MO, Mouritsen OG (2004) Lipids do influence protein function—the hydrophobic matching hypothesis revisited. Biochim Biophys Acta 1666: 205-226. doi: 10.1016/j.bbamem.2004.06.009
|
| [3] |
Dawaliby R, Trubbia C, Delporte C, et al. (2015) Phosphatidylethanolamine Is a Key Regulator of Membrane Fluidity in Eukaryotic Cells. J Biol Chem 291: 3658-3667. doi: 10.1074/jbc.M115.706523
|
| [4] |
Bieberich E (2018) Sphingolipids and lipid rafts: Novel concepts and methods of analysis. Chem Phys Lipids 216: 114-131. doi: 10.1016/j.chemphyslip.2018.08.003
|
| [5] |
Sezgin E, Levental I, Mayor S, et al. (2017) The mistery of membrane organization: composition, regulation and physiological relevance of lipid rafts. Nat Rev Mol Cell Biol 18: 361-374. doi: 10.1038/nrm.2017.16
|
| [6] |
Athenstaedt K, Daum G (1999) Phosphatidic acid, a key intermediate in lipid metabolism. Eur J Biochem 266: 1-16. doi: 10.1046/j.1432-1327.1999.00822.x
|
| [7] |
Bernat P, Gajewska E, Szewczyk R, et al. (2014) Tributyltin (TBT) induces oxidative stress and modifies lipid profile in the filamentous fungus Cunninghamella elegans. Environ Sci Pollut Res 21: 4228-4235. doi: 10.1007/s11356-013-2375-5
|
| [8] |
Voelker DR (2003) New perspectives on the regulation of intermembrane glycerophospholipid traffic. J Lipid Res 44: 441-449. doi: 10.1194/jlr.R200020-JLR200
|
| [9] |
Carman GM, Han GS (2018) Phosphatidate phosphatase regulates membrane phospholipid synthesis via phosphatidylserine synthase. Adv Biol Regul 67: 49-58. doi: 10.1016/j.jbior.2017.08.001
|
| [10] |
Zhang P, Csaki LS, Ronquillo E, et al. (2019) Lipin 2/3 phosphatidic acid phosphatases maintain phospholipid homeostasis to regulate chylomicron synthesis. J Clin Invest 129: 281-295. doi: 10.1172/JCI122595
|
| [11] |
Fonovich T, Magnarelli G (2013) Phosphoinositide and phospholipid phosphorylation and hydrolysis pathways – Organophosphate and organochlorine pesticides effects. Adv Biol Chem 3: 22-35. doi: 10.4236/abc.2013.33A004
|
| [12] | Fonovich de Schroeder TM, Pechén de D'Angelo AM (1991) Dieldrin effects on phospholipid metabolism in Buffo arenarum oocytes. Comp Biochem Physiol 98C: 287-292. |
| [13] | Fonovich de Schroeder TM, Pechén de D'Angelo AM (1995) Dieldrin modifies the hydrolysis of PIP2 and decreases the fertilization rate in Buffo arenarum oocytes. Comp Biochem Physiol 112C: 61-67. |
| [14] |
Nishio K, Sugimoto Y, Fujiwara Y, et al. (1992) Phospholipase C-mediated hydrolysis of phosphatidylcholine is activated by cis-diamminedichloroplatinum (II). J Clin Invest 89: 1622-1628. doi: 10.1172/JCI115758
|
| [15] |
Nakamura Y, Awai K, Masuda T, et al. (2005) A novel phosphatidylcholine-hydrolyzing phospholipase C induced by phosphate starvation in Arabidopsis. J Biol Chem 280: 7469-7476. doi: 10.1074/jbc.M408799200
|
| [16] |
Cruz-Ramírez A, Oropeza-Aburto A, Razo-Hernández F, et al. (2006) Phospholipase DZ2 plays an important role in extraplastidic galactolipid biosynthesis and phosphate recycling in Arabidopsis roots. Proc Natl Acad Sci USA 103: 6765-6770. doi: 10.1073/pnas.0600863103
|
| [17] |
Zavaleta-Pastor M, Sohlenkamp C, Gao JL, et al. (2010) Sinorhizobium meliloti phospholipase C required for lipid remodeling during phosphorus limitation. Proc Natl Acad Sci 107: 302-307. doi: 10.1073/pnas.0912930107
|
| [18] |
Billah MM, Anthes JM (1990) The regulation and cellular functions of phosphatidylcholine hydrolysis. Biochem J 269: 281-291. doi: 10.1042/bj2690281
|
| [19] |
Richmond GS, Smith TK (2011) Phospholipases A1. Int J Mol Sci 12: 588-612. doi: 10.3390/ijms12010588
|
| [20] |
Köhler GA, Brenot A, Haas-Stapleton E, et al. (2006) Phospholipase A2 and Phospholipase B Activities in Fungi. Biochim Biophys Acta 1761: 1391-1399. doi: 10.1016/j.bbalip.2006.09.011
|
| [21] |
Fonovich de Schroeder TM, Pechén de D'Angelo AM (2000) The turnover of phospholipid fatty acyl chains is activated by the insecticide Dieldrin in Buffo arenarum oocytes. J Biochem Molec Toxicol 14: 82-87. doi: 10.1002/(SICI)1099-0461(2000)14:2<82::AID-JBT3>3.0.CO;2-0
|
| [22] |
Wocławek-Potocka I, Rawińska P, Kowalczyk-Zieba I, et al. (2014) Lysophosphatidic Acid (LPA) Signaling in Human and Ruminant Reproductive Tract. Mediators Inflamm 2014: 1-14. doi: 10.1155/2014/649702
|
| [23] |
Ye X, Chun J (2010) Lysophosphatidic Acid (LPA) Signaling in Vertebrate Reproduction. Trends Endocrinol Metab 21: 1-17. doi: 10.1016/j.tem.2009.09.006
|
| [24] |
Kuriyama S, Theveneau E, Benedetto A, et al. (2014) In vivo collective cell migration requires an LPAR2-dependent increase in tissue fluidity. J Cell Biol 206: 113-127. doi: 10.1083/jcb.201402093
|
| [25] |
Jasieniecka-Gazarkiewicz K, Lager I, Carlsson AS, et al. (2017) Acyl-CoA: Lysophosphatidylethanolamine Acyltransferase Activity Regulates Growth of Arabidopsis. Plant Physiol 174: 986-998. doi: 10.1104/pp.17.00391
|
| [26] |
Cullis PR, De Kruijff B (1979) Lipid polymorphism and the functional roles of lipids in biological membranes. Biochim Biophys Acta 559: 399-420. doi: 10.1016/0304-4157(79)90012-1
|
| [27] | Dowhan W, Bogdanov M, Mileykovskaya E (2008) Functional roles of lipids in membranes. Biochemistry of lipids, lipoproteins and membranes Canada: Elsevier, 1-35. |
| [28] |
Ball WB, Neff JK, Gohil VM (2018) The role of non-bilayer phospholipids in mitochondrial structure and function. FEBS Lett 592: 1273-1290. doi: 10.1002/1873-3468.12887
|
| [29] |
Baker CD, Ball WB, Pryce EN, et al. (2016) Specific requirements of nonbilayer phospholipids in mitochondrial respiratory chain function and formation. Mol Biol Cell 27: 2161-2171. doi: 10.1091/mbc.E15-12-0865
|
| [30] |
Gasanov SE, Kim AA, Yaguzhinsky LS, et al. (2018) Non-bilayer Structures in Mitochondrial Membranes Regulate ATP Synthase Activity. Biochim Biophys Acta 1860: 586-599. doi: 10.1016/j.bbamem.2017.11.014
|
| [31] | Fonovich TM, Perez-Coll CS, Fridman O, et al. (2016) Phospholipid changes in Rhinella arenarum embryos under different acclimation conditions to copper. Comp Biochem Physiol Part C 189: 10-16. |
| [32] |
Garay LA, Boundy-Mills KL, Germa JB (2014) Accumulation of High-Value Lipids in Single-Cell Microorganisms: A Mechanistic Approach and Future Perspectives. J Agric Food Chem 67: 2709-2727. doi: 10.1021/jf4042134
|
| [33] |
Welte MA (2015) Expanding roles for lipid droplets. Curr Biol 25: R470-R481. doi: 10.1016/j.cub.2015.04.004
|
| [34] |
Meyers A, Weiskittel TM, Dalhaimer P (2017) Lipid Droplets: Formation to Breakdown. Lipids 52: 465-475. doi: 10.1007/s11745-017-4263-0
|
| [35] |
Olzmann JA, Carvalho P (2019) Dynamics and functions of lipid droplets. Nat Rev Mol Cell Biol 20: 137-155. doi: 10.1038/s41580-018-0085-z
|
| [36] |
Li Z, Thiel K, Thul PJ, et al. (2012) Lipid droplets control the maternal histone supply of Drosophila embryos. Curr Biol 22: 2104-2113. doi: 10.1016/j.cub.2012.09.018
|
| [37] |
Li Z, Johnson MR, Ke Z, et al. (2014) Drosophila lipid droplets buffer the H2Av supply to protect early embryonic development. Curr Biol 24: 1485-1491. doi: 10.1016/j.cub.2014.05.022
|
| [38] |
Huang X, Warren JT, Gilbert LI (2008) New players in the regulation of ecdysone biosynthesis. J Genet Genomics 35: 1-10. doi: 10.1016/S1673-8527(08)60001-6
|
| [39] |
Herms A, Bosch M, Ariotti N, et al. (2013) Cell-to-cell Heterogeneity in Lipid Droplets Suggests a Mechanism to Reduce Lipotoxicity. Curr Biol 23: 1489-1496. doi: 10.1016/j.cub.2013.06.032
|
| [40] |
Grygiel-Górniak B (2014) Peroxisome Proliferator-Activated Receptors and Their Ligands: Nutritional and Clinical Implications. A Review. Nutr J 13: 17-26. doi: 10.1186/1475-2891-13-17
|
| [41] |
Poursharifi P, Madiraju SRM, Prentki M (2017) Monoacylglycerol Signalling and ABHD6 in Health and Disease Diabetes. Obes Metab 19: 76-89. doi: 10.1111/dom.13008
|
| [42] |
Walker OLlS, Holloway AC, Raha S (2019) The role of the endocannabinoid system in female reproductive tissues. J Ovarian Res 12: 3-12. doi: 10.1186/s13048-018-0478-9
|
| [43] |
Fan C, Yan J, Qian Y, et al. (2006) Regulation of Lipoprotein Lipase Expression by Effect of Hawthorn Flavonoids on Peroxisome Proliferator Response Element Pathway. J Pharmacol Sci 100: 51-58. doi: 10.1254/jphs.FP0050748
|
| [44] |
Rotman N, Guex N, Gouranton E, et al. (2013) PPARβ interprets a chromatin signature of pluripotency to promote embryonic differentiation at gastrulation. PLoS One 8: e83300. doi: 10.1371/journal.pone.0083300
|
| [45] | Michalik L, Desvergne B, Dreyer C, et al. (2002) PPAR expression and function during vertebrate development. Int J Dev Biol 46: 105-114. |
| [46] | Fonovich de Schroeder TM (1993) Efecto del Dieldrin sobre la transducción de señales en ovocitos de sapo Bufo arenarum, Hensel. PhD thesis. Pharmacy and Biochemistry Faculty. Buenos Aires University 1-181. |
| [47] | Fonovich de Schroeder TM (1997) Pretreatment ofamphibian oocytes with the organochlorinated pesticide Dieldrin facilitates the formation of the fertilization membrane after insemination. Acta Toxicol Arg 5: 81-83. |
| [48] |
Wozniak KL, Tembo M, Phelps WA, et al. (2018) PLC and IP 3-evoked Ca2+ Release Initiate the Fast Block to Polyspermy in Xenopus laevis Eggs. J Gen Physiol 150: 1239-1248. doi: 10.1085/jgp.201812069
|
| [49] |
Fonovich de Schroeder TM, Pechén de D'Angelo AM (1995) The effect of Dieldrin on Clostridium perfringens posphatidylcholine phospholipase C activity. Pest Biochem Physiol 51: 170-177. doi: 10.1006/pest.1995.1017
|
| [50] |
Carattino MD, Peralta S, Pérez-Coll C, et al. (2004) Effects of Long-Term Exposure to Cu2+ and Cd2+ on the Pentose Phosphate Pathway Dehydrogenase Activities in the Ovary of Adult Bufo Arenarum: Possible Role as Biomarker for Cu2+ Toxicity. Ecotoxicol Environ Saf 57: 311-318. doi: 10.1016/S0147-6513(03)00081-2
|
| [51] | Fonovich de Schroeder TM, Preller AF, Naab F, et al. (2000) Acumulación de Zn en ovocitos de sapo Bufo arenarum: efecto sobre el metabolismo de carbohidratos. Rev Bras Toxicol 13: 55-61. |
| [52] |
Naab F, Volcomirsky M, Burlón A, et al. (2001) Metabolic Alterations Without Metal Accumulation in the Ovary of Adult Bufo Arenarum Females, Observed After Long-Term Exposure to Zn(2+), Followed by Toxicity to Embryos. Arch Environ Contam Toxicol 41: 201-207. doi: 10.1007/s002440010238
|
| [53] |
Fonovich de Schroeder TM (2005) The effect of Zn on glucose 6-phosphate dehydrogenase activity from Bufo arenarum toad ovary and alfalfa plants. Ecotoxicol Environ Saf 60: 123-131. doi: 10.1016/j.ecoenv.2004.07.008
|
| [54] |
Rokitskaya TI, Kotova EA, Agapov II, et al. (2014) Unsaturated lipids protect the integral membrane peptide gramicidin A from singlet oxygen. FEBS Lett 588: 1590-1595. doi: 10.1016/j.febslet.2014.02.046
|
| [55] | Kim SH, Kim BK, Park S, et al. (2019) Phosphatidylcholine extends lifespan via DAF-16 and reduces Amyloid-beta-Induced toxicity in Caenorhabditis elegans. Oxid Med Cell Longev 2019: 2860642. |
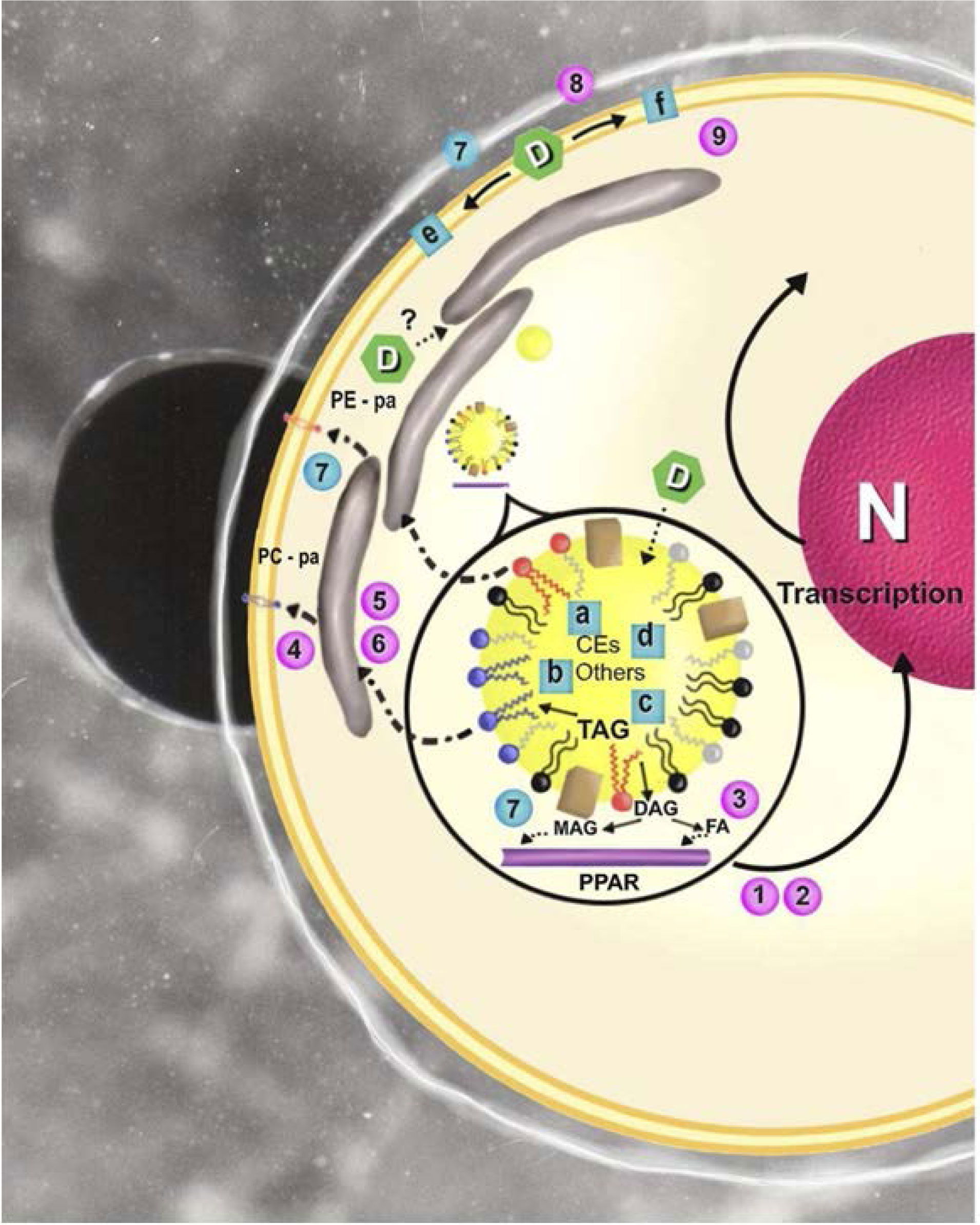
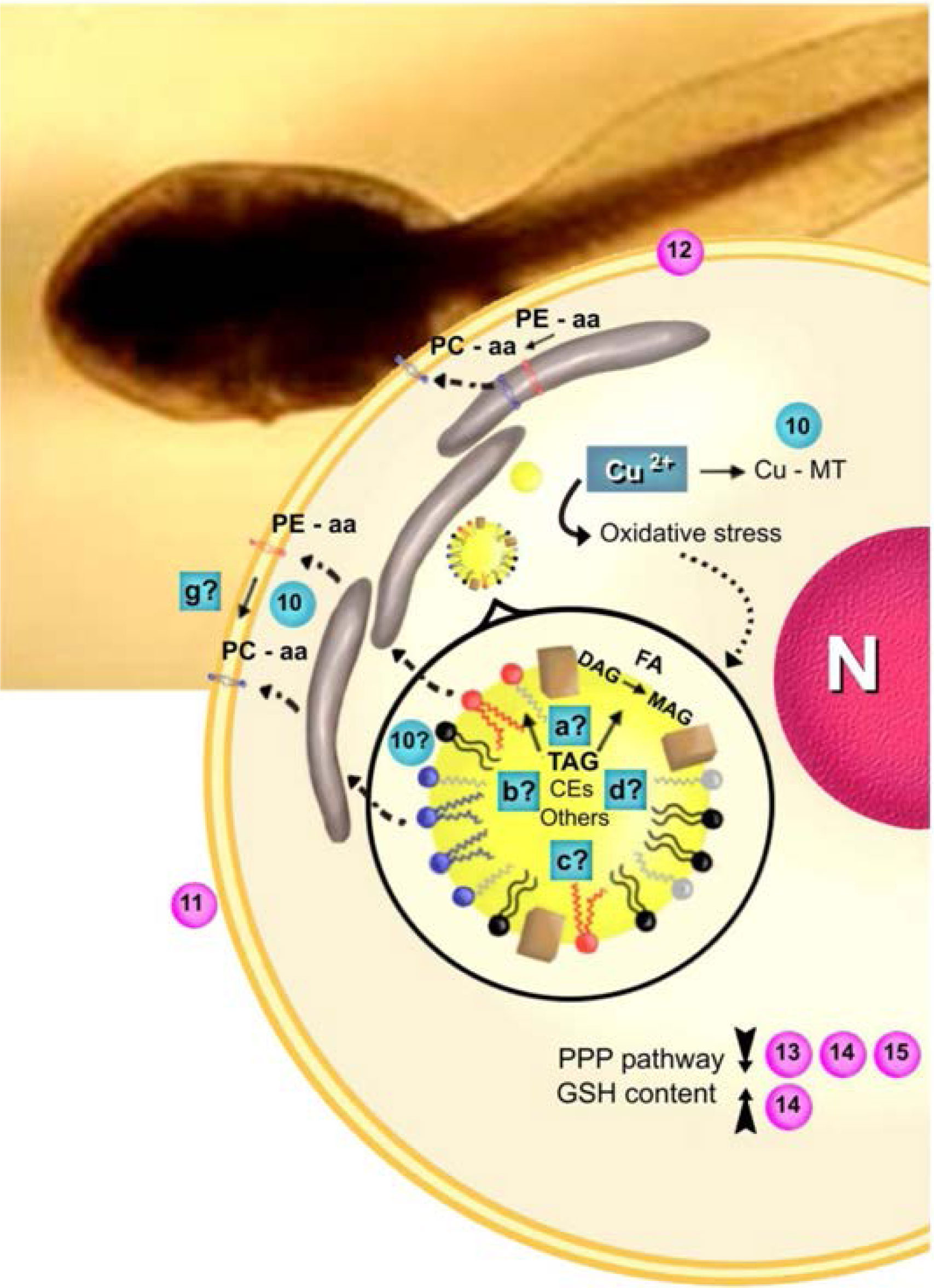
Figures(2)

Teresa M. Fonovich. Phospholipid synthetic and turnover pathways elicited upon exposure to different xenobiotics[J]. AIMS Molecular Science, 2020, 7(3): 211-228. doi: 10.3934/molsci.2020010







 DownLoad:
DownLoad: