Citation: Fumiaki Uchiumi, Makoto Fujikawa, Satoru Miyazaki, Sei-ichi Tanuma. Implication of bidirectional promoters containing duplicated GGAA motifs of mitochondrial function-associated genes[J]. AIMS Molecular Science, 2014, 1(1): 1-26. doi: 10.3934/molsci.2013.1.1

| [1] | Carey MF, Peterson CL, Smale ST (2009) Transcription and preinitiation complex assembly in vitro, In: Carey, M.F., Peterson, C.L., Smale, S.T. Eds., Transcriptional Regulation in Eukaryotes, 2nd Ed, New York: Cold Spring Harbor Laboratory Press, 439-538. |
| [2] |
Scarpulla RC, (2008) Transcriptional paradigms in mammalian mitochondrial biogenesis and function. Physiol Rev 88: 611-638. doi: 10.1152/physrev.00025.2007

|
| [3] |
Campbell CT, Kolesar JE, Kaufman BA, (2012) Mitochondrial transcription factor A regulates mitochondrial transcription initiation, DNA packaging, and genome copy number. Biochim Biophys Acta 1819:921-929. doi: 10.1016/j.bbagrm.2012.03.002
|
| [4] |
Vafai SB, Mootha VK, (2012) Mitochondrial disorders as windows into an ancient organelle. Nature 491: 374-383. doi: 10.1038/nature11707
|
| [5] |
Pagliarini DJ, Calvo SE, Chang B et al. (2008) A mitochondrial protein compendium elucidates complex I disease biology. Cell 134: 112-123. doi: 10.1016/j.cell.2008.06.016
|
| [6] |
Schmidt O, Pfanner N, Meisinger C, (2010) Mitochondrial protein import: from proteomics to functional mechanisms. Nat Rev Mol Cell Biol 11: 655-667. doi: 10.1038/nrm2959
|
| [7] |
Reinders J, Zahedi RP, Pfanner N et al. (2006) Toward the complete yeast mitochondrial proteome: multidimensional separation techniques for mitochondrial proteomics. J. Proteome Res 5: 1543-1554. doi: 10.1021/pr050477f
|
| [8] | Evans MJ, Scarpulla RC (1989) Interaction of nuclear factors with multiple sites in the somatic cytochrome c promoter. Characterization of upstream NRF-1, ATF and intron Sp1 recognition sites. J Biol Chem 264: 14361-14368. |
| [9] |
Scarpulla RC (1997) Nuclear control of respiratory chain expression in mammalian cells. J Bioenerg Biomembr 29: 109-119. doi: 10.1023/A:1022681828846
|
| [10] |
Whatcott CJ, Meyer-Ficca ML, Meyer RG, et al. (2009) A specific isoform of poly(ADP-ribose) glycohydrolase is targeted to the mitochondrial matrix by a N-terminal mitochondrial targeting sequence. Exp Cell Res 315: 3477-3485. doi: 10.1016/j.yexcr.2009.04.005
|
| [11] |
Meyer RG, Meyer-Ficca ML, Jacobson EL, Jacobson MK (2003) Human poly(ADP-ribose) glycohydrolase (PARG) gene and the common promoter sequence it shares with inner mitochondrial membrane translocase 23 (TIM23). Gene 314: 181-190. doi: 10.1016/S0378-1119(03)00738-8
|
| [12] | Uchiumi F, Sakakibara G, Sato J, et al. (2008) Characterization of the promoter region of the human PARG gene and its response to PU.1 during differentiation of HL-60 cells. Genes Cells 13: 1229-1248. |
| [13] |
Uchiumi F, Enokida K, Shiraishi T, et al. (2010) Characterization of the promoter region of the human IGHMBP2 (Smbp-2) gene and its response to TPA in HL-60 cells. Gene 463: 8-17. doi: 10.1016/j.gene.2010.04.014
|
| [14] |
Koc EC, Burkhart W, Blackburn K, et al. (2001) Large subunit of the mammalian mitochondrial ribosome. J Biol Chem 276: 43958-43969. doi: 10.1074/jbc.M106510200
|
| [15] | Uchiumi F, Larsen S, Tanuma S (2013) Biological systems that control transcription of DNA repair-and telomere maintenance-associated genes, In: Chen, C. Ed., DNA repair, Rijeka, Croatia, InTech Open Access Publisher, 309-325. |
| [16] |
Matoba S, Kang JG, Patino WD, et al. (2006) p53 regulates mitochondrial respiration. Science 312: 1650-1653. doi: 10.1126/science.1126863
|
| [17] |
Green DR, Kroemer G (2009) Cytoplasmic functions of the tumor suppressor p53. Nature 458: 1127-1130. doi: 10.1038/nature07986
|
| [18] | Sahin E, DePinho RA (2012) Axis of aging: telomeres, p53 and mitochondria. Nat Rev Mol Cell Biol 13, 397-404. |
| [19] |
Szczesny B, Tann AW, Longley MJ, et al. (2008) Long patch base excision repair in mammalian mitochondrial genomes. J Biol Chem 283: 26349-26356. doi: 10.1074/jbc.M803491200
|
| [20] | Martin SA (2011) Mitochondrial DNA repair, In: Storici, F. Ed., DNA Repair - On the Pathways to Fixing DNA Damage and Errors, Rijeka, Croatia: InTech-Open Access Publisher, 313-338. |
| [21] |
Sementchenko VI, Watson DK (2000) Ets target genes: past, present and future. Oncogene 19: 6533-6548. doi: 10.1038/sj.onc.1204034
|
| [22] |
Baillat D, Laitem C, Leprivier G, et al. (2009) Ets-1 binds cooperatively to the palindromic Ets-binding sites in the p53 promoter. Biochem Biophys Res Commun 378: 213-217. doi: 10.1016/j.bbrc.2008.11.035
|
| [23] |
Molloy-Simard V, St-Laurent JF, Vigneault F, et al. (2012) Altered expression of the poly(ADP-ribosyl)ation enzymes in uveal melanoma and regulation of PARG gene expression by the transcription factor ERM. Invest Ophthamol Vis Sci 53: 6219-6231. doi: 10.1167/iovs.11-8853
|
| [24] |
Uchiumi F, Watanabe T, Tanuma S (2010) Characterization of various promoter regions of the human DNA helicase-encoding genes and identification of duplicated ets (GGAA) motifs as an essential transcription regulatory element. Exp Cell Res 316: 1523-1534. doi: 10.1016/j.yexcr.2010.03.009
|
| [25] |
Uchiumi F, Miyazaki S, Tanuma S (2011) The possible functions of duplicated ets (GGAA) motifs located near transcription start sites of various genes. Cell Mol Life Sci 68: 2039-2051. doi: 10.1007/s00018-011-0674-x
|
| [26] |
Smeitink JA, Loeffen JL, Triepels RH, et al. (1998) Nuclear genes of human complex I of the mitochondrial electron transport chain: state of the art. Hum Mol Genet 7: 1573-1579. doi: 10.1093/hmg/7.10.1573
|
| [27] |
Starker LF, Delgado-Verdugo A, Udelsman R, et al. (2010) Expression and somatic mutations of SDHAF2 (SDH5), a novel endocrine tumor suppressor gene in parathyroid tumors of primary hyperparathyroidism. Endocrine 38: 397-401. doi: 10.1007/s12020-010-9399-0
|
| [28] |
Harris KA, Jones V, Bilbille Y, et al. (2011) YrdC exhibits properties expected of a subunit for a tRNA threonylcarbamoyl transferase. RNA 17: 1678-1687. doi: 10.1261/rna.2592411
|
| [29] |
Yang C, Bolotin E, Jiang T, et al. (2007) Prevalence of the initiator over the TATA box in human and yeast genes and identification of DNA motifs enriched in human TATA-less core promoters. Gene 389: 52-65. doi: 10.1016/j.gene.2006.09.029
|
| [30] | Trinklein ND, Aldred SF, Hartman SJ, et al. (2004) An aboundance of bidirectional promoters in the human genome. Genome Res 14: 62-66. |
| [31] | Yang MQ, Elnitski LL (2008) Diversity of core promoter elements comprising human bidirectional promoters. BMC Genomics 9 (Suppl. 2): S3. |
| [32] |
Collins PJ, Kobayashi Y, Nguyen L, et al. (2007) The ets-related transcription factor GABP directs bidirectional transcription. PLoS Genet 3: e208. doi: 10.1371/journal.pgen.0030208
|
| [33] |
Lin JM, Collins PJ, Trinklein ND, et al. (2007) Transcription factor binding and modified histones in human bidirectional promoters. Genome Res 17: 818-827. doi: 10.1101/gr.5623407
|
| [34] |
Anno YN, Myslinski E, Ngondo-Mbongo RP, et al. (2011) Genome-wide evidence for an essential role of the human Staf/ZNF143 transcription factor in bidirectional transcription. Nucleic Acids Res 39: 3116-3127. doi: 10.1093/nar/gkq1301
|
| [35] |
Orekhova AS, Rubtsov PM (2013) Bidirectional promoters in the transcription of mammalian genomes. Biochem (Moscow) 78: 335-341. doi: 10.1134/S0006297913040020
|
| [36] |
Adachi N, Lieber MR (2002) Bidirctional gene organization: a common architectural feature of the human genome. Cell 109: 807-809. doi: 10.1016/S0092-8674(02)00758-4
|
| [37] |
Wakano C, Byun JS, Di LJ, et al. (2012) The dual lives of bidirectional promoters. Biochim Biophys Acta 1819: 688-693. doi: 10.1016/j.bbagrm.2012.02.006
|
| [38] | Xu C, Chen J, Shen B (2012) The preservation of bidirectional promoter architecture in eukaryotes: what is the driving force? BMC Systems Biol 6 (Suppl. 1): S21. |
| [39] | Uchiumi F, Larsen S, Masumi A, et al. (2013) The putative implications of duplicated GGAA-motifs located in the human interferon regulated genes (ISGs), In: iConcept Ed., Genomics I- Humans, Animals and Plants, Hong Kong: iConcept Press, 87-105. |
| [40] |
Gotea V, Petrykowska HM, Elnitski L (2013) Bidirectional promoters as important drivers for the emergence of species-specific transcripts. PLoS One 8: e57323. doi: 10.1371/journal.pone.0057323
|
| [41] | Yang MQ, Koehly LM, Elinitski LL (2007) Comprehensive annotation of bidirectional promoters identifies co-regulation among breast and ovarian cancer genes. PLoS Comput Biol 3, e72. |
| [42] |
Lin R, Paz S, Hiscott J (2010) Tom70 imports antiviral immunity to the mitochondria. Cell Res 20: 971-973. doi: 10.1038/cr.2010.113
|
| [43] |
Liu XY, Wei B, Shi HX, et al. (2010) Tom70 mediates activation of interferon regulatory factor 3 on mitochondria. Cell Res 20: 994-1011. doi: 10.1038/cr.2010.103
|
| [44] |
Kim SW, Kim JB, Kim JH, et al. (2007) Interferon-gamma-induced expressions of heat shock protein 60 and heat shock protein 10 in C6 astroglioma cells: identification of the signal transducers and activators of transcription 3-binding site in bidirectional promoter. Neuroreport 18: 385-389. doi: 10.1097/WNR.0b013e32801299cc
|
| [45] |
Mauxion F, Chen CY, Séraphin B, et al. (2009) BTG/TOB factors impact deadenylases. Trends Biochem Sci 34: 640-647. doi: 10.1016/j.tibs.2009.07.008
|
| [46] |
Koshiba T (2013) Mitochondrial-mediated antiviral immunity. Biochim Biophys Acta 1833: 225-232. doi: 10.1016/j.bbamcr.2012.03.005
|
| [47] |
Okazaki T, Higuchi M, Gotoh Y (2013) Mitochondrial localization of the antiviral signaling adaptor IPS-1 is important for its induction of caspase activation. Genes Cells 18: 493-501. doi: 10.1111/gtc.12052
|
| [48] | Wei GH, Badis G, Berger MF, et al. Genome-wide analysis of ETS-family DNA-binding in vitro and in vivo. EMBO J 29: 2147-2160. |
| [49] |
Chen YQ, Sengchanthalangsy LL, Hackett A, et al. (2000) F-kappaB p65 (RelA) homodimer uses distinct mechanisms to recognize DNA targets. Structure 8: 419-428. doi: 10.1016/S0969-2126(00)00123-4
|
| [50] |
Wei L, Fan M, Xu L, et al. (2008) Bioinformatic analysis reveals cRel as a regulator of a subset of interferon-stimurated genes. J Interferon Cytokine Res 28: 541-552. doi: 10.1089/jir.2007.0136
|
| [51] |
Nguyen VT, Benveniste EN (2000) Involvement of STAT-1 and Ets family members in interferon-g induction of CD40 transcription in microglia/macrophages. J Biol Chem 275: 23674-23684. doi: 10.1074/jbc.M002482200
|
| [52] | Aittomäki S, Yang J, Scott EW, et al. (2004) Molecular basis of Stat1 and PU.1 cooperation in cytokine-induced Fcgamma receptor 1 promoter activation. Int Immunol 16: 265-274. |
| [53] | Meraro D, Gleit-Kielmanowicz M, Hauser H, et al. (2002) IFN-stimulated gene 15 is synergistically activated through interactions between the myelocyte/lymphocyte-specific transcription factors, PU.1, IFN regulatory factor-8/IFN consensus sequence binding protein, and IFN regulatory factor-4: characterization of a new subtype of IFN-stimulated response element. J Immunol 168: 6224-6231. |
| [54] | Kroeger PE, Morimoto RI (1994) Selection of new HSF1 and HSF2 DNA-binding sites reveals differences in tumor cooperativity. Mol Cell Biol 14: 7592-7603. |
| [55] | Wang G, Qi K, Zhao Y, et al. (2013) Identification of regulatory regions of bidirectional genes in cervical cancer. BMC Med Genet 6 (Suppl. 1): S5. |
| [56] |
Bachman NJ, Wu W, Schmidt TR, et al. (1999) The 5' region of the COX4 gene contains a novel overlapping gene, NOC4. Mamm Genome 10: 506-512. doi: 10.1007/s003359901031
|
| [57] | FitzGerald PC, Shlyakhtenko A, Mir AA, et al. Clustering of DNA sequences in human promoters. Genome Res 14: 1562-1574. |
| [58] | Bellizzi D, Dato S, Cavalcante P, et al. (2005) Characterization of a bidirectional promoter shared between two human genes related to aging: SIRT3 and PSMD13. Genomics 89: 143-150. |
| [59] | Patton J, Block S, Coombs C, et al. (2005) Identification of functional elements in the murine Gabp alpha/ATP synthase coupling factor 6 bi-directional promoter. Gene 369: 35-44. |
| [60] |
Gaston K, Fried M (1994) YY1 is involved in the regulation of the bi-directional promoter of the Surf-1 and Surf-2 genes. FEBS Lett 347: 289-294. doi: 10.1016/0014-5793(94)00567-2
|
| [61] |
Orii KE, Orii KO, Souri M, et al. (1999) Genes for the human mitochondrial trifunctional protein a- and b-subunits are divergently transcribed from a common promoter region. J Biol Chem 274: 8077-8084. doi: 10.1074/jbc.274.12.8077
|
| [62] |
Zanotto E, Häkkinen A, Teku G, et al. (2009) NF-Y influences directionality of transcription from the bidirectional Mrps12/Sarsm promoter in both mouse and human cells. Biochim Biophys Acta 1789: 432-442. doi: 10.1016/j.bbagrm.2009.05.001
|
| [63] | Merika M, Thanos D (2011) Enhanceosomes. Curr Opin Genet Dev 11: 205-208. |
| [64] |
Sahin E, Colla S, Liesa M, et al. (2011) Telomere dysfunction induces metabolic and mitochondrial compromise. Nature 470: 359-365. doi: 10.1038/nature09787
|
| [65] | O'Sullivan RJ, Karlseder J (2010) Telomeres: protecting chromosomes against genome instability. Nat Rev Mol Cell Biol 11: 171-181. |
| [66] |
Yu BP, Chung HY (2006) Adaptive mechanisms to oxidative stress during aging. Mech Ageing Dev 127: 436-443. doi: 10.1016/j.mad.2006.01.023
|
| [67] |
Wolff S, Dillin A (2013) Beneficial miscommunication. Nature 497: 442-443. doi: 10.1038/497442a
|
| [68] |
Houtkooper RH, Mouchiroud L, Ryu D, et al. (2013) Mitochondrial protein imbalance as a conserved longevity mechanism. Nature 497: 451-457. doi: 10.1038/nature12188
|
| [69] |
Schmacher B (2009) Transcription-blocking DNA damage in aging: a mechanism for hormesis. Bioessays 31: 1347-1356. doi: 10.1002/bies.200900107
|
| [70] |
Calabrese EJ, Mattson MP, Calabrase V (2010) Resveratrol commonly displays hormesis: Occurrence and biomedical significance. Hum Exp Toxicol 29: 980-1015. doi: 10.1177/0960327110383625
|
| [71] |
Howitz KT, Bitterman KJ, Cohen HY, et al. (2003) Small molecule activators of sirtuins extend Saccharomyces cerevisiae life span. Nature 425: 191-196. doi: 10.1038/nature01960
|
| [72] |
Wood JG, Rogina B, Lavul S, et al. (2004) Sirtuin activators mimic caloric restriction and delay aging in metazoans. Nature 430: 686-689. doi: 10.1038/nature02789
|
| [73] |
Stefani M, Markus MA, Lin RC, et al. (2007) The effect of resveratrol on a cell model of human aging. Ann NY Acad Sci 1114: 407-418. doi: 10.1196/annals.1396.001
|
| [74] | Kroemer G (1997) Mitochondrial implication in apoptosis. Towards an endosymbiont hypothesis of apoptosis evolution. Cell Death Differ 4: 443-546. |
| [75] |
Tait SW, Green DR (2010) Mitochondria and cell death: outer membrane permeabilization and beyond. Nat Rev Mol Cell Biol 11: 621-632. doi: 10.1038/nrm2952
|
| [76] | Estaquier J, Vallette F, Vayssiere JL, et al. (2012) The mitochondrial pathways of apoptosis, In: Scatena, R., Bottoni, P., Giardina B. Eds., Advances in Mitochondrial Medicine, Dordrecht, Germany: Springer Science+Business Media B.V., 157-183. |
| [77] |
Vandenabeele P, Galluzzi L, Berghe TV (2010) Molecular mechanisms of necroptosis: an ordered cellular explosion. Nat Rev Mol Cell Biol 11: 700-714. doi: 10.1038/nrm2970
|
| [78] |
Galluzzi L, Kepp O, Kroemer G (2012) Mitochondria: master regulators of danger signaling. Nat Rev Mol Cell Biol 13: 780-788. doi: 10.1038/nrm3479
|
| [79] |
Buchakjian MR, Kornbluth S (2010) The engine driving the ship: metabolic steering of cell proliferation and death. Nat Rev Mol Cell Biol 11: 715-727. doi: 10.1038/nrm2972
|
| [80] |
Gut P (2013) The nexus of chromatin regulation and intermediary metabolism. Nature 502: 489-498. doi: 10.1038/nature12752
|
| [81] |
Pearce EL, Poffenberger MC, Chang CH, et al. (2013) Fueling Immunity: Insights into metabolism and lymphocyte function. Science 342: 1242454. doi: 10.1126/science.1242454
|
| [82] |
Tan-Wong SM, Zaugg JB, Camblong J, et al. (2012) Gene loops enhance transcriptional directionality. Science 338: 671-675. doi: 10.1126/science.1224350
|
| [83] |
Ansari A, Hampsey M (2005) A role for the CPF 3'-end processing machinery in RNAP II-dependent gene looping. Genes Dev 19: 2969-2978. doi: 10.1101/gad.1362305
|
| [84] |
Hampsey M (2012) A new direction for gene loops. Science 338: 624-625. doi: 10.1126/science.1230576
|
| [85] |
Lykke-Andersen S, Mapendano CK, Jensen TH (2011) An ending is a new beginning. Cell Cycle 10: 863-865. doi: 10.4161/cc.10.6.14931
|
| [86] |
Perkins KJ, Lusic M, Mitar I, et al. (2008) Transcription-dependent gene looping of the HIV-1 provirus is dictated by recognition of pre-mRNA processing signals. Mol Cell 29: 56-68. doi: 10.1016/j.molcel.2007.11.030
|
| [87] |
Barrett LW, Fletcher S, Wilton SD (2012) Regulation of eukaryotic gene expression by the untranslated gene regions and other non-coding elements. Cell Mol Life Sci 69: 3613-3634. doi: 10.1007/s00018-012-0990-9
|
| [88] |
McCue AD, Nuthikattu S, Reeder S.H, et al. (2012) Gene expression and stress response mediated by the epigenetic regulation of a transposable element small RNA. PLoS Genet 8: e1002474. doi: 10.1371/journal.pgen.1002474
|
| [89] |
Kapitonov VV, Jurka J (2005) RAG1 core and V(D)J recombination signal sequences were derived from Transib transposons. PLoS Biol 3: 181. doi: 10.1371/journal.pbio.0030181
|
| [90] |
Acquaviva L, Székvölgyi L, Dichtl B, et al. (2013) The COMPASS subunit Spp1 links histone methylation to initiation of meiotic recombination. Science 339: 215-218. doi: 10.1126/science.1225739
|
| [91] |
Core LT, Waterfall JJ, Lis JT (2008) Nascent RNA sequencing reveals widespread pausing and divergent initiation at human promoters. Science 322: 1845-1848. doi: 10.1126/science.1162228
|
| [92] |
Seila AC, Calabrese JM, Levine SS, et al. (2008) Divergent transcription from active promoters. Science 322: 1849-1851. doi: 10.1126/science.1162253
|
| [93] |
Preker P, Nielsen J, Kammler S, et al. (2008) RNA exosome depletion reveals transcription upstream of active human promoters. Science 322: 1851-1854. doi: 10.1126/science.1164096
|
| [94] |
Almada AE, Wu X, Kriz AJ (2013) Promoter directionality is controlled by U1 snRNP and polyadenylation signals. Nature 499: 360-363. doi: 10.1038/nature12349
|
| [95] |
Ntini E, Järvelin AI, Bornholdt J et al. (2013) Polyadenylation site-induced decay of upstream transcripts enforces promoter directionality. Nat Struct Mol Biol 20: 923-928. doi: 10.1038/nsmb.2640
|
| [96] |
Zhang L, Ding Q, Wang P, et al. (2013) An upstream promoter element blocks the reverse transcription of the mouse insulin-degrading enzyme gene. Biochem Biophys Res Commun 430: 26-31. doi: 10.1016/j.bbrc.2012.11.052
|
| [97] |
Szklarczyk R, Huynen MA (2010) Mosaic origin of the mitochondrial proteome. Proteomics 10: 4012-4024. doi: 10.1002/pmic.201000329
|
| [98] |
Yang D, Oyaizu Y, Oyaizu H, et al. (1985) Mitochondrial origins. Proc Natl Acad Sci USA 82: 4443-4447. doi: 10.1073/pnas.82.13.4443
|
| [99] |
Viale AM, Arakaki AK (1994) The chaperone connection to the origins of the eukaryotic organelles. FEBS Lett 341: 146-151. doi: 10.1016/0014-5793(94)80446-X
|
| [100] |
Andersson SG, Zomorodipour A, Andersson JO, et al. (1998) The genome sequence of Rickettsia prowazekii and the origin of mitochondria. Nature 396: 133-143. doi: 10.1038/24094
|
| [101] |
Dunning Hotopp JC (2011) Horizontal gene transfer between bacteria and animals. Trends Genet 27: 157-163. doi: 10.1016/j.tig.2011.01.005
|
| [102] |
Schönknecht G, Chen WH, Ternes CM, et al. (2013) Gene transfer from bacteria and archaea facilitated evolution of an extremophilic eukaryote. Science 339: 1207-1210. doi: 10.1126/science.1231707
|
| [103] | Martin WF (2013) Early evolution without a tree of life. Biol Direct 6: 36. |
| [104] |
Rocha EPC (2013) With a little help from prokaryotes. Science 339: 1154-1155. doi: 10.1126/science.1234938
|
| [105] |
ENCODE Project Consortium (2012) An integrated encyclopedia of DNA elements in the human genome. Nature 489: 57-74. doi: 10.1038/nature11247
|
| [106] | Scatena R (2012) Mitochondria and cancer, In: Scatena, R., Bottoni, P., Giardina B. Eds., Advances in Mitochondrial Medicine, Dordrecht, Germany: Springer Science+Business Media B.V., 287-308. |
| [107] |
Schultze A, Harris AL (2012) How cancer metabolism is turned for proliferation and vulnerable to disruption. Nature 491: 364-373. doi: 10.1038/nature11706
|
| [108] | Camara AK, Bienengraeber M, Stowe DF (2011) Mitochondrial approaches to protect against cardiac ischemia and reperfusion injury. Front Physiol 2: 1-34. |
| [109] | Griffiths EJ (2012) Mitochondria and Heart Disease, In: Scatena, R., Bottoni, P., Giardina B. Eds., Advances in Mitochondrial Medicine, Dordrecht, Germany: Springer Science Business Media B.V., 249-267. |
| [110] | Swerdlow LE, Swerdlow RH (2012) Mitochondria in nerodegeneration, In: Scatena, R., Bottoni, P., Giardina B. Eds., Advances in Mitochondrial Medicine, Dordrecht, Germany: Springer Science Business Media B.V., 269-286. |
| [111] |
Petit A, Kawarai T, Paitel E, et al. (2005) Wild-type PINK1 prevents basal and induced neuronal apoptosis, a protective effect abrogated by Parkinson-disease-related mutations. J Biol Chem 280: 34025-34032. doi: 10.1074/jbc.M505143200
|
| [112] |
Dagda RK, Chu CT (2009) Mitochondrial quality control: insights on how Parkinson's disease related genes PINK1, parkin, and Omi/HtrA2 interact to maintain mitochondrial homeostasis. J Bioenerg Biomembr 41: 473-479. doi: 10.1007/s10863-009-9255-1
|
| [113] |
Hertz NT, Berthet A, Sos ML, et al. (2013) A neo-substrate that amplifies catalytic activity of Parkinson's-disease-related kinase PINK1. Cell 154: 737-747. doi: 10.1016/j.cell.2013.07.030
|
| [114] |
Courchet J, Lewis TL, Lee S, et al. (2013) Terminal axon branching is regulated by the LKB1-NUAK1 kinase pathway via presynaptic mitochondrial capture. Cell 153: 1510-1525. doi: 10.1016/j.cell.2013.05.021
|
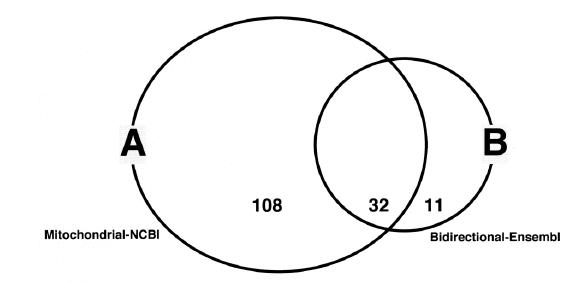
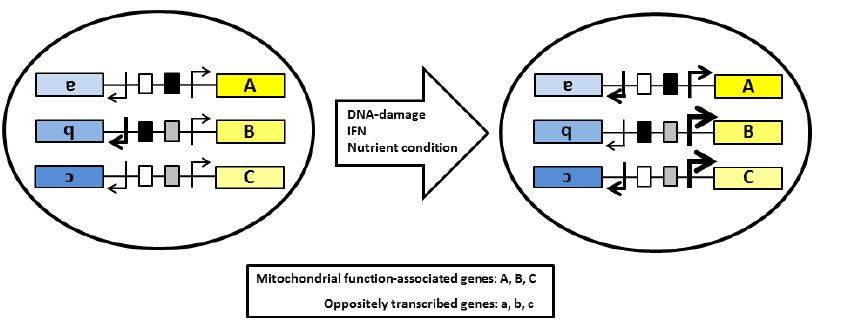
Figures(2) / Tables(4)

Fumiaki Uchiumi, Makoto Fujikawa, Satoru Miyazaki, Sei-ichi Tanuma. Implication of bidirectional promoters containing duplicated GGAA motifs of mitochondrial function-associated genes[J]. AIMS Molecular Science, 2014, 1(1): 1-26. doi: 10.3934/molsci.2013.1.1







 DownLoad:
DownLoad: