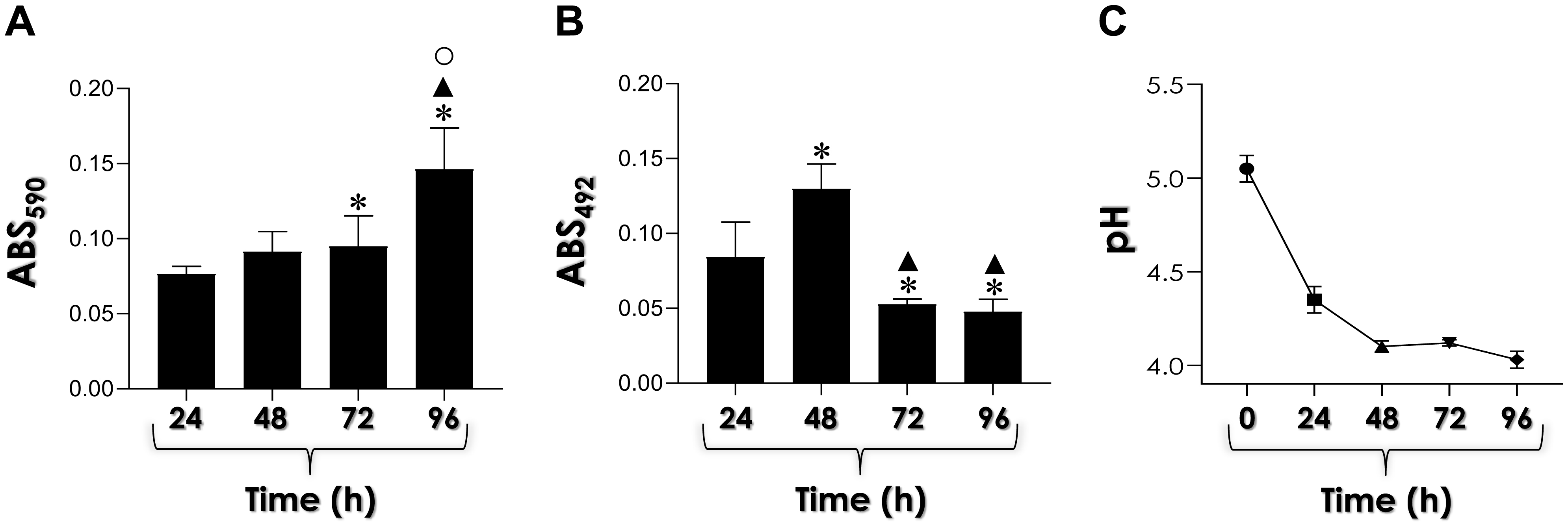
Candida haemulonii is an emerging, opportunistic, and multidrug-resistant fungal pathogen. Recently, our group reported the ability of C. haemulonii to form biofilm and secrete aspartic-type peptidases (Saps). Herein, we investigated the correlation between Saps production and biofilm formation along C. haemulonii growth in yeast carbon base medium supplemented with albumin (a Sap-inducing condition) and in the presence of the classical Sap inhibitor pepstatin A. Under these conditions, the biofilm biomass increased on a polystyrene surface, reaching its maximum at 96 h, while maximum biofilm viability was detected at 48 h. The release of Saps during biofilm formation showed an inverse trend, with the highest enzymatic activity measured after 24 h. In the presence of pepstatin A, a significant reduction in biofilm parameters (biomass and viability), as well as in albumin consumption by biofilm-forming cells was detected. These findings underscore the importance of Saps during the biofilm development in C. haemulonii.
Citation: Joice Cavalcanti Lima, Lívia de Souza Ramos, Pedro Fernandes Barbosa, Iuri Casemiro Barcellos, Marta Helena Branquinha, André Luis Souza dos Santos. Biofilm production by the multidrug-resistant fungus Candida haemulonii is affected by aspartic peptidase inhibitor[J]. AIMS Microbiology, 2025, 11(1): 228-241. doi: 10.3934/microbiol.2025012
Candida haemulonii is an emerging, opportunistic, and multidrug-resistant fungal pathogen. Recently, our group reported the ability of C. haemulonii to form biofilm and secrete aspartic-type peptidases (Saps). Herein, we investigated the correlation between Saps production and biofilm formation along C. haemulonii growth in yeast carbon base medium supplemented with albumin (a Sap-inducing condition) and in the presence of the classical Sap inhibitor pepstatin A. Under these conditions, the biofilm biomass increased on a polystyrene surface, reaching its maximum at 96 h, while maximum biofilm viability was detected at 48 h. The release of Saps during biofilm formation showed an inverse trend, with the highest enzymatic activity measured after 24 h. In the presence of pepstatin A, a significant reduction in biofilm parameters (biomass and viability), as well as in albumin consumption by biofilm-forming cells was detected. These findings underscore the importance of Saps during the biofilm development in C. haemulonii.
Amprenavir
Brain Heart Infusion
Bovine Serum Albumin
Darunavir
Human Immunodeficiency Virus
Lopinavir
Nelfinavir
Phosphate-Buffered Saline
1,10-Phenanthroline
Phenylmethylsulfonyl Fluoride
Roswell Park Memorial Institute
Ritonavir
Secreted Aspartic Peptidases
Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis
2,3-Bis(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide (Tetrazolium Salt)
Yeast Carbon Base
Yeast Extract-Peptone-Dextrose

| [1] |
Jong AW, Al-Obaid K, Mohd Tap R, et al. (2023) Candida khanbhai sp. nov., a new clinically relevant yeast within the Candida haemulonii species complex. Med Mycol 61: myad009. https://doi.org/10.1093/mmy/myad009 
|
| [2] |
Cendejas-Bueno E, Kolecka A, Alastruey-Izquierdo A, et al. (2012) Reclassification of the Candida haemulonii complex as Candida haemulonii (C. haemulonii group I), C. duobushaemulonii sp. nov. (C. haemulonii group II), and C. haemulonii var. vulnera var. nov.: three multiresistant human pathogenic yeasts. J Clin Microbiol 50: 3641-3651. https://doi.org/10.1128/JCM.02248-12
|
| [3] |
Ramos LS, Figueiredo-Carvalho MHG, Silva LN, et al. (2022) The threat called Candida haemulonii species complex in Rio de Janeiro–Brazil: focus on antifungal resistance and virulence attributes. J Fungi 8: 574. https://doi.org/10.3390/jof8060574
|
| [4] |
Ramos LS, Oliveira SSC, Souto XM, et al. (2017) Planktonic growth and biofilm formation profiles in Candida haemulonii species complex. Med Mycol 55: 785-789. https://doi.org/10.1093/mmy/myx005
|
| [5] |
Malinovská Z, Čonková E, Váczi P (2023) Biofilm formation in medically important Candida species. J Fungi 10: 955. https://doi.org/10.3390/jof9100955
|
| [6] |
Monika S, Małgorzata B, Zbigniew O (2017) Contribution of aspartic proteases in Candida virulence. Protease inhibitors against Candida infections. Curr Protein Pept Sci 18: 1050-1062. https://doi.org/10.2174/1389203717666160809155749
|
| [7] |
Staniszewska M (2020) Virulence factors in Candida species. Curr Protein Pept Sci 21: 313-323. https://doi.org/10.2174/1389203720666190722152415
|
| [8] |
Ramos LS, Oliveira SSC, Braga-Silva LA, et al. (2020) Secreted aspartyl peptidases by the emerging, opportunistic and multidrug-resistant fungal pathogens comprising the Candida haemulonii complex. Fungal Biol 124: 700-707. https://doi.org/10.1016/j.funbio.2020.04.001
|
| [9] |
Dutton LC, Jenkinson HF, Lamont RJ, et al. (2016) Role of Candida albicans secreted aspartyl protease Sap9 in interkingdom biofilm formation. Pathog Dis 74: ftw005. https://doi.org/10.1093/femspd/ftw005
|
| [10] |
Winter MB, Salcedo EC, Lohse MB, et al. (2016) Global identification of biofilm-specific proteolysis in Candida albicans. mBio 7: e01514-16. https://doi.org/10.1128/mBio.01514-16
|
| [11] |
Ramos LS, Figueiredo-Carvalho MH, Barbedo LS, et al. (2015) Candida haemulonii complex: species identification and antifungal susceptibility profiles of clinical isolates from Brazil. J Antimicrob Chemother 70: 111-115. https://doi.org/10.1093/jac/dku321
|
| [12] |
White TC, Agabian N (1995) Candida albicans secreted aspartyl proteinases: isoenzyme pattern is determined by cell type, and levels are determined by environmental factors. J Bacteriol 177: 5215-5221. https://doi.org/10.1128/jb.177.18.5215-5221.1995
|
| [13] |
Braga-Silva LA, Mogami SS, Valle RS, et al. (2010) Multiple effects of amprenavir against Candida albicans. FEMS Yeast Res 10: 221-224. https://doi.org/10.1111/j.1567-1364.2009.00595.x
|
| [14] |
Peeters E, Nelis HJ, Coenye T (2008) Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J Microbiol Methods 72: 157-165. https://doi.org/10.1016/j.mimet.2007.11.010
|
| [15] |
Ramos LS, Mello TP, Branquinha MH, et al. (2020) Biofilm formed by Candida haemulonii species complex: structural analysis and extracellular matrix composition. J Fungi 6: 46. https://doi.org/10.3390/jof6020046
|
| [16] |
Lowry OH, Rosebrough NJ, Farr AL, et al. (1951) Protein measurement with the Folin phenol reagent. J Biol Chem 193: 265-275.
|
| [17] |
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685. https://doi.org/10.1038/227680a0
|
| [18] |
Santos ALS, Braga-Silva LA, Gonçalves DS, et al. (2021) Repositioning lopinavir, an HIV protease inhibitor, as a promising antifungal drug: lessons learned from Candida albicans – in silico, in vitro and in vivo approaches. J Fungi 7: 424. https://doi.org/10.3390/jof7060424
|
| [19] |
Pereira R, Dos Santos Fontenelle RO, de Brito EHS, et al. (2021) Biofilm of Candida albicans: formation, regulation and resistance. J Appl Microbiol 131: 11-22. https://doi.org/10.1111/jam.14949
|
| [20] |
Serrano-Fujarte I, López-Romero E, Reyna-López GE, et al. (2015) Influence of culture media on biofilm formation by Candida species and response of sessile cells to antifungals and oxidative stress. Biomed Res Int 2015: 783639. https://doi.org/10.1155/2015/783639
|
| [21] |
Tan Y, Leonhard M, Ma S, et al. (2016) Influence of culture conditions for clinically isolated non-albicans Candida biofilm formation. J Microbiol Methods 130: 123-128. https://doi.org/10.1016/j.mimet.2016.09.011
|
| [22] |
Hirota K, Yumoto H, Sapaar B, et al. (2017) Pathogenic factors in Candida biofilm-related infectious diseases. J Appl Microbiol 122: 321-330. https://doi.org/10.1111/jam.13330
|
| [23] |
Kadry AA, El-Ganiny AM, El-Baz AM (2018) Relationship between Sap prevalence and biofilm formation among resistant clinical isolates of Candida albicans. African Health Sci 18: 1166-1174. https://doi.org/10.4314/ahs.v18i4.37
|
| [24] |
Kim JS, Lee KT, Bahn YS (2023) Secreted aspartyl protease 3 regulated by the Ras/cAMP/PKA pathway promotes the virulence of Candida auris. Front Cell Infect Microbiol 13: 1257897. https://doi.org/10.3389/fcimb.2023.1257897
|
| [25] |
Muñoz JF, Gade L, Chow NA, et al. (2018) Genomic insights into multidrug-resistance, mating and virulence in Candida auris and related emerging species. Nat Commun 9: 5346. https://doi.org/10.1038/s41467-018-07779-6
|
| [26] |
Fenley JC, de Barros PP, Carmo PHFD, et al. (2022) Repurposing HIV protease inhibitors atazanavir and darunavir as antifungal treatments against Candida albicans infections: an in vitro and in vivo study. Curr Issues Mol Biol 44: 5379-5389. https://doi.org/10.3390/cimb44110364
|
| [27] |
Consolaro ME, Gasparetto A, Svidzinski TI, et al. (2006) Effect of pepstatin A on the virulence factors of Candida albicans strains isolated from vaginal environment of patients in three different clinical conditions. Mycopathologia 162: 75-82. https://doi.org/10.1007/s11046-006-0026-9
|
| [28] |
Souto XM, Branquinha MH, Santos ALS (2019) Chymotrypsin- and trypsin-like activities secreted by the multidrug-resistant yeasts forming the Candida haemulonii complex. Anais Acad Bras Ciênc 91: e20180735. https://doi.org/10.1590/00013765201920180735
|



Figures(4)

Joice Cavalcanti Lima, Lívia de Souza Ramos, Pedro Fernandes Barbosa, Iuri Casemiro Barcellos, Marta Helena Branquinha, André Luis Souza dos Santos. Biofilm production by the multidrug-resistant fungus Candida haemulonii is affected by aspartic peptidase inhibitor[J]. AIMS Microbiology, 2025, 11(1): 228-241. doi: 10.3934/microbiol.2025012







 DownLoad:
DownLoad: