Citation: Arnab Bhowmik, Mara Cloutier, Emily Ball, Mary Ann Bruns. Underexplored microbial metabolisms for enhanced nutrient recycling in agricultural soils[J]. AIMS Microbiology, 2017, 3(4): 826-845. doi: 10.3934/microbiol.2017.4.826

| [1] |
Phelps J, Carrasco LR, Webb EL, et al. (2013) Agricultural intensification escalates future conservation costs. P Natl Acad Sci USA 110: 7601–7606. doi: 10.1073/pnas.1220070110

|
| [2] |
Lal R (2004) Soil carbon sequestration impacts on global climate change and food security. Science 304: 1623–1627. doi: 10.1126/science.1097396
|
| [3] |
West TO, Post WM (2002) Soil organic carbon sequestration rates by tillage and crop rotation. Soil Sci Soc Am J 66: 1930–1946. doi: 10.2136/sssaj2002.1930
|
| [4] |
Galloway JN, Dentener FJ, Capone DG, et al. (2004) Nitrogen cycles: past, present, and future. Biogeochemistry 70: 153–226. doi: 10.1007/s10533-004-0370-0
|
| [5] | Inselsbacher E (2009) The fate of inorganic nitrogen fertilizers in agricultural soils: University of Vienna, 179. |
| [6] |
Tilman DG, Cassman KG, Matson PA, et al. (2002) Agricultural sustainability and intensive production practices. Nature 418: 671–677. doi: 10.1038/nature01014
|
| [7] |
Gomiero T (2016) Soil degradation, land scarcity and food security: Reviewing a complex challenge. Sustainability 8: 281. doi: 10.3390/su8030281
|
| [8] |
Habig J, Swanepoel C (2015) Effects of conservation agriculture and fertilization on soil microbial diversity and activity. Environments 2: 358–384. doi: 10.3390/environments2030358
|
| [9] |
Conant RT, Easter M, Paustian K, et al. (2007) Impacts of periodic tillage on soil C stocks: A synthesis. Soil Till Res 95: 1–10. doi: 10.1016/j.still.2006.12.006
|
| [10] |
Drinkwater LE, Wagoner P, Sarrantonio M (1998) Legume-based cropping systems have reduced carbon and nitrogen losses. Nature 396: 262–265. doi: 10.1038/24376
|
| [11] |
McDaniel MD, Tiemann LK, Grandy AS (2014) Does agricultural crop diversity enhance soil microbial biomass and organic matter dynamics? A meta‐analysis. Ecol Appl 24: 560–570. doi: 10.1890/13-0616.1
|
| [12] | Bhowmik A, Fortuna A, Cihacek LJ, et al. (2017) Potential carbon sequestration and nitrogen cycling in long-term organic management systems. Renew Agr Food Syst: 1–13. |
| [13] |
Morriën E, Hannula SE, Snoek LB, et al. (2017) Soil networks become more connected and take up more carbon as nature restoration progresses. Nat Commun 8: 14349. doi: 10.1038/ncomms14349
|
| [14] | Paul EA, Morris SJ, Bohm S (2001) The determination of soil C pool sizes and turnover rates: biophysical fractionation and tracers, In: Lal R, Kimble JM, Follett RF, et al., Editors, Assessment Methods for Soil Carbon, Lewis Pubilshers, 193–206. |
| [15] | Alexander M (1977) Introduction to soil microbiology, John Wiley & Sons. |
| [16] |
Krebs HA (1941) Carbon dioxide assimilation in heterotrophic organisms. Nature 147: 560–563. doi: 10.1038/147560a0
|
| [17] |
Reicosky DC, Archer DW (2007) Moldboard plow tillage depth and short-term carbon dioxide release. Soil Till Res 94: 109–121. doi: 10.1016/j.still.2006.07.004
|
| [18] |
Wu X, Ge T, Yuan H, et al. (2014) Changes in bacterial CO2 fixation with depth in agricultural soils. Appl Micobiol Biot 98: 2309–2319. doi: 10.1007/s00253-013-5179-0
|
| [19] |
Roslev P, Larsen MB, Jørgensen D, et al. (2004) Use of heterotrophic CO2 assimilation as a measure of metabolic activity in planktonic and sessile bacteria. J Microbiol Meth 59: 381–393. doi: 10.1016/j.mimet.2004.08.002
|
| [20] |
Miltner A, Richnow H, Kopinke F, et al. (2004) Assimilation of CO2 by soil microorganisms and transformation into soil organic matter. Org Geochem 35: 1015–1024. doi: 10.1016/j.orggeochem.2004.05.001
|
| [21] |
Miltner A, Richnow H, Kopinke F, et al. (2005) Incorporation of carbon originating from CO2 into different compounds of soil microbial biomass and soil organic matter. Isot Environ Healt S 41: 135–140. doi: 10.1080/10256010412331314265
|
| [22] | Bernalier A, Willems A, Leclerc M, et al. (1996) Ruminococcus hydrogenotrophicus sp. nov., a new H2/CO2-utilizing acetogenic bacterium isolated from human feces. Arch Microbiol 166: 176–183. |
| [23] |
Barker HA, Kamen MD, Haas V (1945) Carbon dioxide utilization in the synthesis of acetic and butyric acids by Butyribacterium rettgeri. P Natl Acad Sci USA 31: 355–360. doi: 10.1073/pnas.31.11.355
|
| [24] |
Jones SW, Fast AG, Carlson ED, et al. (2016) CO2 fixation by anaerobic non-photosynthetic mixotrophy for improved carbon conversion. Nat Commun 7: 12800. doi: 10.1038/ncomms12800
|
| [25] | Hobbie JE, Hobbie EA (2013) Microbes in nature are limited by carbon and energy: the starving-survival lifestyle in soil and consequences for estimating microbial rates. Front Microbiol 4. |
| [26] |
Meredith LK, Rao D, Bosak T, et al. (2014) Consumption of atmospheric hydrogen during the life cycle of soil‐dwelling Actinobacteria. Env Microbiol Rep 6: 226–238. doi: 10.1111/1758-2229.12116
|
| [27] |
Greening C, Constant P, Hards K, et al. (2015) Atmospheric hydrogen scavenging: from enzymes to ecosystems. Appl Environ Microb 81: 1190–1199. doi: 10.1128/AEM.03364-14
|
| [28] |
Rhee TS, Brenninkmeijer CAM, Röckmann T (2006) The overwhelming role of soils in the global atmospheric hydrogen cycle. Atmos Chem Phys 6: 1611–1625. doi: 10.5194/acp-6-1611-2006
|
| [29] |
Maimaiti J, Zhang Y, Yang J, et al. (2007) Isolation and characterization of hydrogen‐oxidizing bacteria induced following exposure of soil to hydrogen gas and their impact on plant growth. Environ Microbiol 9: 435–444. doi: 10.1111/j.1462-2920.2006.01155.x
|
| [30] |
Piché-Choquette S, Tremblay J, Tringe SG, et al. (2016) H2-saturation of high affinity H2-oxidizing bacteria alters the ecological niche of soil microorganisms unevenly among taxonomic groups. PeerJ 4: e1782. doi: 10.7717/peerj.1782
|
| [31] | La FJS, Focht DD (1983) Conservation in soil of H2 liberated from N2 fixation by Hup-nodules. Appl Environ Microb 46: 304–311. |
| [32] |
Witty JF (1991) Microelectrode measurements of hydrogen concentrations and gradients in legume nodules. J Exp Bot 42: 765–771. doi: 10.1093/jxb/42.6.765
|
| [33] |
Dong Z, Layzell DB (2001) H2 oxidation, O2 uptake and CO2 fixation in hydrogen treated soils. Plant Soil 229: 1–12. doi: 10.1023/A:1004810017490
|
| [34] |
Tiso M, Schechter AN (2015) Nitrate reduction to nitrite, nitric oxide and ammonia by gut bacteria under physiological conditions. PloS One 10: e0119712. doi: 10.1371/journal.pone.0119712
|
| [35] |
Greening C, Berney M, Hards K, et al. (2014) A soil actinobacterium scavenges atmospheric H2 using two membrane-associated, oxygen-dependent [NiFe] hydrogenases. P Natl Acad Sci USA 111: 4257–4261. doi: 10.1073/pnas.1320586111
|
| [36] | Liot Q, Constant P (2016) Breathing air to save energy–new insights into the ecophysiological role of high‐affinity [NiFe]‐hydrogenase in Streptomyces avermitilis. MicrobiologyOpen 5: 47–59. |
| [37] |
Braun K, Gottschalk G (1981) Effect of molecular hydrogen and carbon dioxide on chemo-organotrophic growth of Acetobacterium woodii and Clostridium aceticum. Arch Microbiol 128: 294–298. doi: 10.1007/BF00422533
|
| [38] | Kellum R, Drake HL (1984) Effects of cultivation gas phase on hydrogenase of the acetogen Clostridium thermoaceticum. J Bacteriol 160: 466–469. |
| [39] |
Krumholz LR, Bradstock P, Sheik CS, et al. (2015) Syntrophic growth of Desulfovibrio alaskensis requires genes for H2 and formate metabolism as well as those for flagellum and biofilm formation. Appl Environ Microb 81: 2339–2348. doi: 10.1128/AEM.03358-14
|
| [40] |
Schmitt S, Hanselmann A, Wollschläger U, et al. (2009) Investigation of parameters controlling the soil sink of atmospheric molecular hydrogen. Tellus B 61: 416–423. doi: 10.1111/j.1600-0889.2008.00402.x
|
| [41] | Smith‐Downey NV, Randerson JT, Eiler JM (2006) Temperature and moisture dependence of soil H2 uptake measured in the laboratory. Geophys Res Lett 33. |
| [42] |
Constant P, Poissant L, Villemur R (2009) Tropospheric H2 budget and the response of its soil uptake under the changing environment. Sci Total Environ 407: 1809–1823. doi: 10.1016/j.scitotenv.2008.10.064
|
| [43] | Team CW (2014) Climate change 2014: Synthesis report. Contribution of working groups I, II and III to the fifth assessment of the Intergovernmental Panel on Climate Change, Geneva, 151. |
| [44] | Zumft WG (1997) Cell biology and molecular basis of denitrification. Microbiol Mol Biol Rev 61: 533–616. |
| [45] |
Dandie CE, Burton DL, Zebarth BJ, et al. (2007) Analysis of denitrification genes and comparison of nosZ, cnorB and 16S rDNA from culturable denitrifying bacteria in potato cropping systems. Syst Appl Microbiol 30: 128–138. doi: 10.1016/j.syapm.2006.05.002
|
| [46] |
Sanford RA, Wagner DD, Wu Q, et al. (2012) Unexpected nondenitrifier nitrous oxide reductase gene diversity and abundance in soils. P Natl Acad Sci USA 109: 19709–19714. doi: 10.1073/pnas.1211238109
|
| [47] |
Henry S, Bru D, Stres B, et al. (2006) Quantitative detection of the nosZ gene, encoding nitrous oxide reductase, and comparison of the abundances of 16S rRNA, narG, nirK, and nosZ genes in soils. Appl Environ Microb 72: 5181–5189. doi: 10.1128/AEM.00231-06
|
| [48] | Smith CJ, Nedwell DB, Dong LF, et al. (2007) Diversity and abundance of nitrate reductase genes (narG and napA), nitrite reductase genes (nirS and nrfA), and their transcripts in estuarine sediments. Appl Environ Microb 73: 3612–3622. |
| [49] | Kamp A, Høgslund S, Risgaard-Petersen N, et al. (2015) Nitrate storage and dissimilatory nitrate reduction by eukaryotic microbes. Front Microbiol 6. |
| [50] |
Simon J (2002) Enzymology and bioenergetics of respiratory nitrite ammonification. FEMS Microbiol Rev 26: 285–309. doi: 10.1111/j.1574-6976.2002.tb00616.x
|
| [51] |
Strohm TO, Griffin B, Zumft WG, et al. (2007) Growth yields in bacterial denitrification and nitrate ammonification. Appl Environ Microb 73: 1420–1424. doi: 10.1128/AEM.02508-06
|
| [52] |
Kern M, Simon J (2008) Characterization of the NapGH quinol dehydrogenase complex involved in Wolinella succinogenes nitrate respiration. Mol Microbiol 69: 1137–1152. doi: 10.1111/j.1365-2958.2008.06361.x
|
| [53] |
Cole J (1996) Nitrate reduction to ammonia by enteric bacteria: redundancy, or a strategy for survival during oxygen starvation? FEMS Microbiol Lett 136: 1–11. doi: 10.1111/j.1574-6968.1996.tb08017.x
|
| [54] | Tiedje JM (1988) Ecology of denitrification and dissimilatory nitrate reduction to ammonium. Biol Anaerob Microorgan 717: 179–244. |
| [55] |
Fazzolari É, Nicolardot B, Germon JC (1998) Simultaneous effects of increasing levels of glucose and oxygen partial pressures on denitrification and dissimilatory nitrate reduction to ammonium in repacked soil cores. Eur J Soil Biol 34: 47–52. doi: 10.1016/S1164-5563(99)80006-5
|
| [56] | Tiedje JM, Sexstone AJ, Myrold DD, et al. (1982) Denitrification: ecological niches, competition and survival. Anton Leeuw 48: 569–583. |
| [57] |
Simon J, Klotz MG (2013) Diversity and evolution of bioenergetic systems involved in microbial nitrogen compound transformations. BBA-Bioenergetics 1827: 114–135. doi: 10.1016/j.bbabio.2012.07.005
|
| [58] |
Cunha CA, Macieira S, Dias JM, et al. (2003) Cytochrome c nitrite reductase from Desulfovibrio desulfuricans ATCC 27774 the relevance of the two calcium sites in the structure of the catalytic subunit (nrfA). J Biol Chem 278: 17455–17465. doi: 10.1074/jbc.M211777200
|
| [59] |
Atkinson SJ, Mowat CG, Reid GA, et al. (2007) An octaheme c‐type cytochrome from Shewanella oneidensis can reduce nitrite and hydroxylamine. FEBS Lett 581: 3805–3808. doi: 10.1016/j.febslet.2007.07.005
|
| [60] |
Doyle RAS, Marritt SJ, Gwyer JD, et al. (2013) Contrasting catalytic profiles of multiheme nitrite reductases containing CxxCK heme-binding motifs. J Biol Inorg Chem 18: 655–667. doi: 10.1007/s00775-013-1011-7
|
| [61] |
Mohan SB, Schmid M, Jetten M, et al. (2004) Detection and widespread distribution of the nrfA gene encoding nitrite reduction to ammonia, a short circuit in the biological nitrogen cycle that competes with denitrification. FEMS Microbiol Ecol 49: 433–443. doi: 10.1016/j.femsec.2004.04.012
|
| [62] |
Kraft B, Strous M, Tegetmeyer HE (2011) Microbial nitrate respiration-genes, enzymes and environmental distribution. J Biotechnol 155: 104–117. doi: 10.1016/j.jbiotec.2010.12.025
|
| [63] | Song B, Lisa JA, Tobias CR (2014) Linking DNRA community structure and activity in a shallow lagoonal estuarine system. Front Microbiol 5. |
| [64] |
Welsh A, Chee-Sanford JC, Connor LM, et al. (2014) Refined NrfA phylogeny improves PCR-based nrfA gene detection. Appl Environ Microb 80: 2110–2119. doi: 10.1128/AEM.03443-13
|
| [65] |
Tatti E, Goyer C, Zebarth BJ, et al. (2017) Over-winter dynamics of soil bacterial denitrifiers and nitrite ammonifiers influenced by crop residues with different carbon to nitrogen ratios. Appl Soil Ecol 110: 53–64. doi: 10.1016/j.apsoil.2016.10.014
|
| [66] |
Bhowmik A, Fortuna A, Cihacek LJ, et al. (2016) Use of biological indicators of soil health to estimate reactive nitrogen dynamics in long-term organic vegetable and pasture systems. Soil Biol Biochem 103: 308–319. doi: 10.1016/j.soilbio.2016.09.004
|
| [67] |
Stremińska MA, Felgate H, Rowley G, et al. (2012) Nitrous oxide production in soil isolates of nitrate‐ammonifying bacteria. Env Microbiol Rep 4: 66–71. doi: 10.1111/j.1758-2229.2011.00302.x
|
| [68] | Firestone MK, Davidson EA (1989) Microbiological basis of NO and N2O production and consumption in soil, In: Exchange of trace gases between terrestrial ecosystems and the atmosphere, 7–21. |
| [69] |
Zhu X, Burger M, Doane TA, et al. (2013) Ammonia oxidation pathways and nitrifier denitrification are significant sources of N2O and NO under low oxygen availability. P Natl Acad Sci USA 110: 6328–6333. doi: 10.1073/pnas.1219993110
|
| [70] |
Rütting T, Boeckx P, Müller C, et al. (2011) Assessment of the importance of dissimilatory nitrate reduction to ammonium for the terrestrial nitrogen cycle. Biogeosciences 8: 1779–1791. doi: 10.5194/bg-8-1779-2011
|
| [71] |
Matheson FE, Nguyen ML, Cooper AB, et al. (2002) Fate of 15N-nitrate in unplanted, planted and harvested riparian wetland soil microcosms. Ecol Eng 19: 249–264. doi: 10.1016/S0925-8574(02)00093-9
|
| [72] |
Clay DE, Molina JAE, Clapp CE, et al. (1990) Soil tillage impact on the diurnal redox-potential cycle. Soil Sci Soc Am J 54: 516–521. doi: 10.2136/sssaj1990.03615995005400020038x
|
| [73] |
Yin SX, Chen D, Chen LM, et al. (2002) Dissimilatory nitrate reduction to ammonium and responsible microorganisms in two Chinese and Australian paddy soils. Soil Biol Biochem 34: 1131–1137. doi: 10.1016/S0038-0717(02)00049-4
|
| [74] |
Pett-Ridge J, Silver WL, Firestone MK (2006) Redox fluctuations frame microbial community impacts on N-cycling rates in a humid tropical forest soil. Biogeochemistry 81: 95–110. doi: 10.1007/s10533-006-9032-8
|
| [75] | Em VDB, Van DU, Abbas B, et al. (2015) Enrichment of DNRA bacteria in a continuous culture. Int Soc Micobiol Ecol 9: 2153–2161. |
| [76] | Jones CM, Graf DRH, Bru D, et al. (2013) The unaccounted yet abundant nitrous oxide-reducing microbial community: a potential nitrous oxide sink. Int Soc Micobiol Ecol 7: 417–426. |
| [77] | Harter J, Krause H, Schuettler S, et al. (2014) Linking N2O emissions from biochar-amended soil to the structure and function of the N-cycling microbial community. Int Soc Micobiol Ecol 8: 660–674. |
| [78] |
Gao J, Xie Y, Jin H, et al. (2016) Nitrous oxide emission and denitrifier abundance in two agricultural soils amended with crop residues and urea in the north China plain. PloS One 11: e0154773. doi: 10.1371/journal.pone.0154773
|
| [79] |
Maeda K, Spor A, Edel-Hermann V, et al. (2015) N2O production, a widespread trait in fungi. Sci Rep 5: 9697. doi: 10.1038/srep09697
|
| [80] | Gorfer M, Klaubauf S, Berger H, et al. (2014) The fungal contribution to the nitrogen cycle in agricultural soils, In: Marco D, Editor, Metagenomics of the Microbial Nitrogen Cycle: Theory, Methods and Applications, 209–225. |
| [81] | Orellana LH, Rodriguez-R LM, Higgins S, et al. (2014) Detecting nitrous oxide reductase (nosZ) genes in soil metagenomes: method development and implications for the nitrogen cycle. Mbio 5: e01193-14. |
| [82] | Sanford RA, Cole JR, Tiedje JM (2002) Characterization and description of Anaeromyxobacter dehalogenans gen. nov., sp. nov., an aryl-halorespiring facultative anaerobic myxobacterium. Appl Environ Microb 68: 893–900. |
| [83] |
Yoon S, Nissen S, Park D, et al. (2016) Nitrous oxide reduction kinetics distinguish bacteria harboring clade I NosZ from those harboring clade II NosZ. Appl Environ Microb 82: 3793–3800. doi: 10.1128/AEM.00409-16
|
| [84] |
Coyotzi S, Doxey AC, Clark ID, et al. (2017) Agricultural soil denitrifiers possess extensive nitrite reductase gene diversity. Environ Microbiol 19: 1189–1208. doi: 10.1111/1462-2920.13643
|
| [85] |
Thompson KA, Bent E, Abalos D, et al. (2016) Soil microbial communities as potential regulators of in situ N2O fluxes in annual and perennial cropping systems. Soil Biol Biochem 103: 262–273. doi: 10.1016/j.soilbio.2016.08.030
|
| [86] |
Bru D, Sarr A, Philippot L (2007) Relative abundances of proteobacterial membrane-bound and periplasmic nitrate reductases in selected environments. Appl Environ Microb 73: 5971–5974. doi: 10.1128/AEM.00643-07
|
| [87] |
Frey SD, Elliott ET, Paustian K (1999) Bacterial and fungal abundance and biomass in conventional and no-tillage agroecosystems along two climatic gradients. Soil Biol Biochem 31: 573–585. doi: 10.1016/S0038-0717(98)00161-8
|
| [88] |
Helgason BL, Walley FL, Germida JJ (2009) Fungal and bacterial abundance in long-term no-till and intensive-till soils of the Northern Great Plains. Soil Sci Soc Am J 73: 120–127. doi: 10.2136/sssaj2007.0392
|
| [89] |
McGonigle TP, Miller MH (1996) Mycorrhizae, phosphorus absorption, and yield of maize in response to tillage. Soil Sci Soc Am J 60: 1856–1861. doi: 10.2136/sssaj1996.03615995006000060034x
|
| [90] |
Kabir Z (2005) Tillage or no-tillage: impact on mycorrhizae. Can J Plant Sci 85: 23–29. doi: 10.4141/P03-160
|
| [91] |
Sipilä TP, Yrjälä K, Alakukku L, et al. (2012) Cross-site soil microbial communities under tillage regimes: fungistasis and microbial biomarkers. Appl Environ Microb 78: 8191–8201. doi: 10.1128/AEM.02005-12
|
| [92] |
Slot JC, Hibbett DS (2007) Horizontal transfer of a nitrate assimilation gene cluster and ecological transitions in fungi: a phylogenetic study. PloS One 2: e1097. doi: 10.1371/journal.pone.0001097
|
| [93] | Marzluf GA (1997) Genetic regulation of nitrogen metabolism in the fungi. Microbiol Mol Biol Rev 61: 17–32. |
| [94] |
Nygren CMR, Eberhardt U, Karlsson M, et al. (2008) Growth on nitrate and occurrence of nitrate reductase‐encoding genes in a phylogenetically diverse range of ectomycorrhizal fungi. New Phytol 180: 875–889. doi: 10.1111/j.1469-8137.2008.02618.x
|
| [95] | Gorfer M, Blumhoff M, Klaubauf S, et al. (2011) Community profiling and gene expression of fungal assimilatory nitrate reductases in agricultural soil. Int Soc Micobiol Ecol 5: 1771–1783. |
| [96] |
Klaubauf S, Inselsbacher E, Zechmeister-Boltenstern S, et al. (2010) Molecular diversity of fungal communities in agricultural soils from Lower Austria. Fungal Divers 44: 65–75. doi: 10.1007/s13225-010-0053-1
|
| [97] | Hankinson O, Cove DJ (1974) Regulation of the pentose phosphate pathway in the fungus Aspergillus nidulans. The effect of growth with nitrate. J Biol Chem 249: 2344–2353. |
| [98] | Hankinson O (1974) Mutants of the pentose phosphate pathway in Aspergillus nidulans. J Bacteriol 117: 1121–1130. |
| [99] |
Schinko T, Berger H, Lee W, et al. (2010) Transcriptome analysis of nitrate assimilation in Aspergillus nidulans reveals connections to nitric oxide metabolism. Mol Microbiol 78: 720–738. doi: 10.1111/j.1365-2958.2010.07363.x
|
| [100] |
Raynaud X, Nunan N (2014) Spatial ecology of bacteria at the microscale in soil. PloS One 9: e87217. doi: 10.1371/journal.pone.0087217
|
| [101] |
Lu W, Zhang H, Min J, et al. (2015) Dissimilatory nitrate reduction to ammonium in a soil under greenhouse vegetable cultivation as affected by organic amendments. J Soils Sed 15: 1169–1177. doi: 10.1007/s11368-015-1089-3
|
| [102] |
DeCatanzaro JB, Beauchamp EG, Drury CF (1987) Denitrification vs dissimilatory nitrate reduction in soil with alfalfa, straw, glucose and sulfide treatments. Soil Biol Biochem 19: 583–587. doi: 10.1016/0038-0717(87)90102-7
|
| [103] |
Miltner A, Kopinke F, Kindler R, et al. (2005) Non-phototrophic CO2 fixation by soil microorganisms. Plant Soil 269: 193–203. doi: 10.1007/s11104-004-0483-1
|
| [104] |
Harter J, Weigold P, El-Hadidi M, et al. (2016) Soil biochar amendment shapes the composition of N2O-reducing microbial communities. Sci Total Environ 562: 379–390. doi: 10.1016/j.scitotenv.2016.03.220
|
| [105] |
Inselsbacher E, Hinko-Najera UN, Stange FC, et al. (2010) Short-term competition between crop plants and soil microbes for inorganic N fertilizer. Soil Biol Biochem 42: 360–372. doi: 10.1016/j.soilbio.2009.11.019
|
| [106] |
Bronick CJ, Lal R (2005) Soil structure and management: a review. Geoderma 124: 3–22. doi: 10.1016/j.geoderma.2004.03.005
|
| [107] |
Cole JA (1978) The rapid accumulation of large quantities of ammonia during nitrite reduction by Escherichia coli. FEMS Microbiol Lett 4: 327–329. doi: 10.1111/j.1574-6968.1978.tb02891.x
|
| [108] |
Burns LC, Stevens RJ, Laughlin RJ (1995) Determination of the simultaneous production and consumption of soil nitrite using 15N. Soil Biol Biochem 27: 839–844. doi: 10.1016/0038-0717(94)00219-Q
|
| [109] |
Stevens RJ, Laughlin RJ (1998) Measurement of nitrous oxide and di-nitrogen emissions from agricultural soils. Nutr Cycl Agroecosys 52: 131–139. doi: 10.1023/A:1009715807023
|
| [110] |
Venterea RT (2007) Nitrite‐driven nitrous oxide production under aerobic soil conditions: kinetics and biochemical controls. Global Change Biol 13: 1798–1809. doi: 10.1111/j.1365-2486.2007.01389.x
|
| [111] |
Hansel CM, Fendorf S, Jardine PM, et al. (2008) Changes in bacterial and archaeal community structure and functional diversity along a geochemically variable soil profile. Appl Environ Microb 74: 1620. doi: 10.1128/AEM.01787-07
|
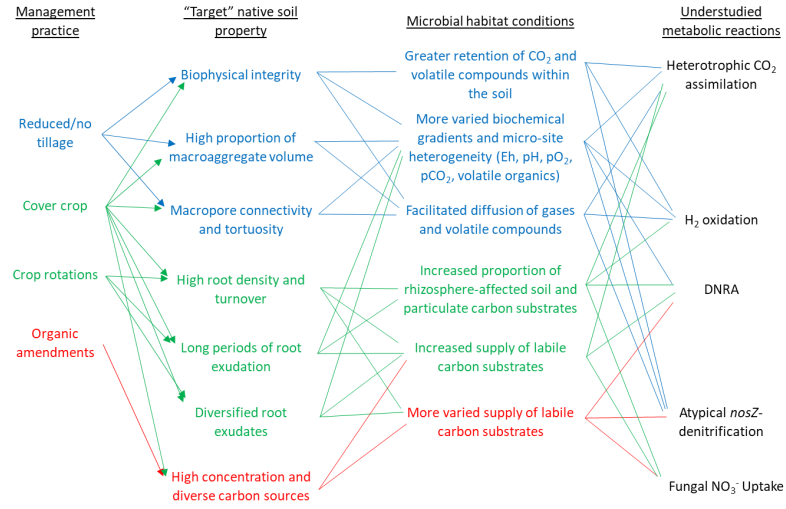
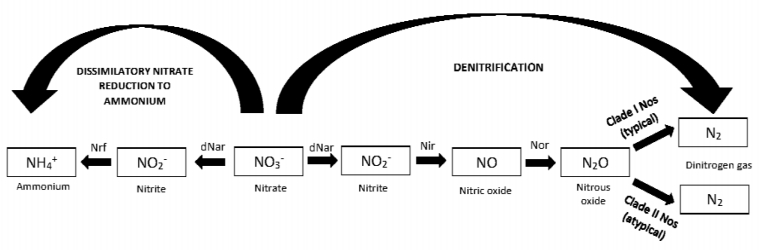
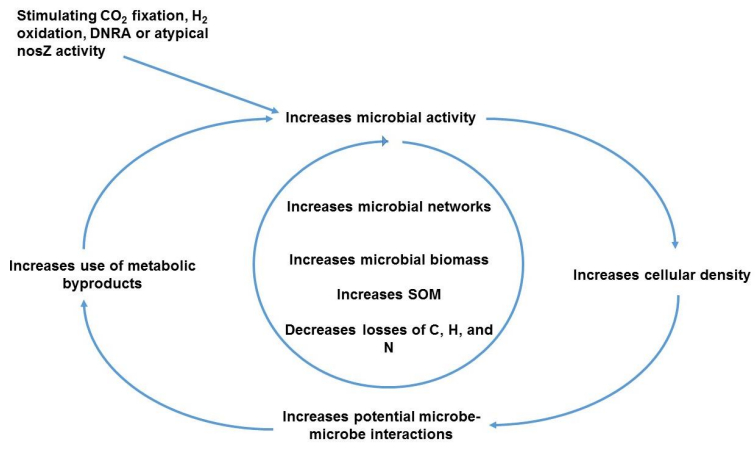
Figures(3)

Arnab Bhowmik, Mara Cloutier, Emily Ball, Mary Ann Bruns. Underexplored microbial metabolisms for enhanced nutrient recycling in agricultural soils[J]. AIMS Microbiology, 2017, 3(4): 826-845. doi: 10.3934/microbiol.2017.4.826







 DownLoad:
DownLoad: