Cooperation is an indispensable behavior in biological systems. In the prisoner's dilemma, due to the individual's selfish psychology, the defector is in the dominant position finally, which results in a social dilemma. In this paper, we discuss the replicator dynamics of the prisoner's dilemma with penalty and mutation. We first discuss the equilibria and stability of the prisoner's dilemma with a penalty. Then, the critical delay of the bifurcation with the payoff delay as the bifurcation parameter is obtained. In addition, considering the case of player mutation based on penalty, we analyze the two-delay system containing payoff delay and mutation delay and find the critical delay of Hopf bifurcation. Theoretical analysis and numerical simulations show that cooperative and defective strategies coexist when only a penalty is added. The larger the penalty is, the more players tend to cooperate, and the critical time delay of the time-delay system decreases with the increase in penalty. The addition of mutation has little effect on the strategy chosen by players. The two-time delay also causes oscillation.
Citation: Yifei Wang, Xinzhu Meng. Evolutionary game dynamics of cooperation in prisoner's dilemma with time delay[J]. Mathematical Biosciences and Engineering, 2023, 20(3): 5024-5042. doi: 10.3934/mbe.2023233
Cooperation is an indispensable behavior in biological systems. In the prisoner's dilemma, due to the individual's selfish psychology, the defector is in the dominant position finally, which results in a social dilemma. In this paper, we discuss the replicator dynamics of the prisoner's dilemma with penalty and mutation. We first discuss the equilibria and stability of the prisoner's dilemma with a penalty. Then, the critical delay of the bifurcation with the payoff delay as the bifurcation parameter is obtained. In addition, considering the case of player mutation based on penalty, we analyze the two-delay system containing payoff delay and mutation delay and find the critical delay of Hopf bifurcation. Theoretical analysis and numerical simulations show that cooperative and defective strategies coexist when only a penalty is added. The larger the penalty is, the more players tend to cooperate, and the critical time delay of the time-delay system decreases with the increase in penalty. The addition of mutation has little effect on the strategy chosen by players. The two-time delay also causes oscillation.

| [1] | J. W. Weibull, Evolutionary Game Theory, MIT press, 1997. |
| [2] | J. M. Smith, Evolution and the Theory of Games, Cambridge university press, 1982. |
| [3] | J. Tanimoto, Fundamentals of Rvolutionary Game Theory and its Applications, Springer Japan, 2015. |
| [4] | J. M. Smith, G. R. Price, The logic of animal conflict, Nature, 246 (1973), 15–18. https://doi.org/10.1038/246015a0 |
| [5] | R. Axelrod, W. D. Hamilton, The evolution of cooperation, Science, 211 (1981), 1390–1396. https://www.science.org/doi/abs/10.1126/science.7466396 |
| [6] |
W. Zhang, Y. S. Li, C. Xu, P. M. Hui, Cooperative behavior and phase transitions in co-evolving stag hunt game, Phys. A, 443 (2016), 161–169. https://doi.org/10.1016/j.physa.2015.09.047 doi: 10.1016/j.physa.2015.09.047

|
| [7] |
J. M. Du, Z. R. Wu, Evolutionary dynamics of cooperation in dynamic networked systems with active striving mechanism, Appl. Math. Comput., 430 (2022), 127295. https://doi.org/10.1016/j.amc.2022.127295 doi: 10.1016/j.amc.2022.127295
|
| [8] |
R. Chiong, M. Kirley, Random mobility and the evolution of cooperation in spatial N-player iterated Prisoner's Dilemma games, Phys. A, 391 (2012), 3915–3923. https://doi.org/10.1016/j.physa.2012.03.010 doi: 10.1016/j.physa.2012.03.010
|
| [9] |
J. L. Chen, X. W. Liu, H. Z. Wang, J. Yang, The disconnection-reconnection-elite mechanism enhances cooperation of evolutionary game on lattice, Chaos Solitons Fractals, 157 (2022), 111897. https://doi.org/10.1016/j.chaos.2022.111897 doi: 10.1016/j.chaos.2022.111897
|
| [10] |
J. X. Pi, G. H. Yang, H. Yang, Evolutionary dynamics of cooperation in N-person snowdrift games with peer punishment and individual disguise, Phys. A, 592 (2022), 126839. https://doi.org/10.1016/j.physa.2021.126839 doi: 10.1016/j.physa.2021.126839
|
| [11] |
P. C. Zhu, H. Guo, H. L. Zhang, Y. Han, Z. Wang, C. Chu, The role of punishment in the spatial public goods game, Nonlinear Dyn., 102 (2020), 2959–2968. https://doi.org/10.1007/s11071-020-05965-0 doi: 10.1007/s11071-020-05965-0
|
| [12] |
P. Catalán, J. M. Seoane, M. A. Sanjuán, Mutation-selection equilibrium in finite populations playing a Hawk–Dove game, Commun. Nonlinear Sci. Numer. Simul., 25 (2015), 66–73. https://doi.org/10.1016/j.cnsns.2015.01.012 doi: 10.1016/j.cnsns.2015.01.012
|
| [13] |
T. Nagatani, G. Ichinose, K. I. Tainaka, Metapopulation model for rock–paper–scissors game: mutation affects paradoxical impacts, J. Theor. Biol., 450 (2018), 22–29. https://doi.org/10.1016/j.jtbi.2018.04.005 doi: 10.1016/j.jtbi.2018.04.005
|
| [14] |
T. A. Wettergren, Replicator dynamics of an N-player snowdrift game with delayed payoffs, Appl. Math. Comput., 404 (2021), 126204. https://doi.org/10.1016/j.amc.2021.126204 doi: 10.1016/j.amc.2021.126204
|
| [15] |
J. Alboszta, J. Miekisz, Stability of evolutionarily stable strategies in discrete replicator dynamics with time delay, J. Theor. Biol., 231 (2004), 175–179. https://doi.org/10.1016/j.jtbi.2004.06.012 doi: 10.1016/j.jtbi.2004.06.012
|
| [16] |
J. Miekisz, S. Wesołowski, Stochasticity and time delays in evolutionary games, Dyn. Games Appl., 1 (2011), 440–448. https://doi.org/10.1007/s13235-011-0028-1 doi: 10.1007/s13235-011-0028-1
|
| [17] |
J. Burridge, Y. Gao, Y. Mao, Delayed response in the Hawk Dove game, Eur. Phys. J. B, 90 (2017), 1–6. https://doi.org/10.1140/epjb/e2016-70471-1 doi: 10.1140/epjb/e2016-70471-1
|
| [18] |
S. Mittal, A. Mukhopadhyay, S. Chakraborty, Evolutionary dynamics of the delayed replicator-mutator equation: Limit cycle and cooperation, Phys. Rev. E, 101 (2020), 042410. https://doi.org/10.1103/PhysRevE.101.042410 doi: 10.1103/PhysRevE.101.042410
|
| [19] |
L. M. Hu, X. L. Qiu, Stability analysis of game models with fixed and stochastic delays, Appl. Math. Comput., 435 (2022), 127473. https://doi.org/10.1016/j.amc.2022.127473 doi: 10.1016/j.amc.2022.127473
|
| [20] |
J. Tanimoto, H. Sagara, Relationship between dilemma occurrence and the existence of a weakly dominant strategy in a two-player symmetric game, Biosystems, 90 (2007), 105–114. https://doi.org/10.1016/j.biosystems.2006.07.005 doi: 10.1016/j.biosystems.2006.07.005
|
| [21] |
Z. Wang, S. Kokubo, M. Jusup, J. Tanimoto, Universal scaling for the dilemma strength in evolutionary games, Phys. Life Rev., 14 (2015), 1–30. https://doi.org/10.1016/j.plrev.2015.04.033 doi: 10.1016/j.plrev.2015.04.033
|
| [22] |
H. Ito, J. Tanimoto, Scaling the phase-planes of social dilemma strengths shows game-class changes in the five rules governing the evolution of cooperation, R. Soc. Open Sci., 5 (2018), 181085. https://doi.org/10.1098/rsos.181085 doi: 10.1098/rsos.181085
|
| [23] |
M. R. Arefin, K. M. A. Kabir, M. Jusup, et al. Social efficiency deficit deciphers social dilemmas, Sci. Rep., 10 (2020), 16092. https://doi.org/10.1038/s41598-020-72971-y doi: 10.1038/s41598-020-72971-y
|
| [24] | J. Tanimoto, Sociophysics Approach to Epidemics, Singapore, Springer, 2021. |
| [25] |
M. Doebeli, C. Hauert, Models of cooperation based on the Prisoner's Dilemma and the Snowdrift game, Ecol. Lett., 8 (2005), 748–766. https://doi.org/10.1111/j.1461-0248.2005.00773.x doi: 10.1111/j.1461-0248.2005.00773.x
|
| [26] | S. Fan, A new extracting formula and a new distinguishing means on the one variable cubic equation, Nat. Sci. J. Hainan Teach. Coll, 2 (1989), 91–98. |
| [27] |
S. Zhang, R. Clark, Y. K. Huang, Frequency-dependent strategy selection in a hunting game with a finite population, Appl. Math. Comput., 382 (2020), 125355. https://doi.org/10.1016/j.amc.2020.125355 doi: 10.1016/j.amc.2020.125355
|
| [28] |
F. Yan, X. Chen, Z. Qiu, A. Szolnoki, Cooperator driven oscillation in a time-delayed feedback-evolving game, New J. Phys., 23 (2021), 053017. https://dx.doi.org/10.1088/1367-2630/abf205 doi: 10.1088/1367-2630/abf205
|
| [29] | D. Pais, C. H. Caicedo-Nunez, N. E. Leonard, Hopf bifurcations and limit cycles in evolutionary network dynamics, SIAM J. Appl. Dyn. Syst., 11 (2012). 1754–1784. https://doi.org/10.1137/120878537 |
| [30] |
A. Szolnoki, M. Perc, Decelerated invasion and waning-moon patterns in public goods games with delayed distribution, Phys. Rev. E, 87 (2013), 054801. https://doi.org/10.1103/PhysRevE.87.054801 doi: 10.1103/PhysRevE.87.054801
|
| [31] |
X. W. Wang, S. Nie, L. L. Jiang, B. H. Wang, S. M. Chen, Role of delay-based reward in the spatial cooperation, Phys. A, 456 (2017), 153–158. https://doi.org/10.1016/j.physa.2016.08.014 doi: 10.1016/j.physa.2016.08.014
|
| [32] |
D. Helbing, A. Szolnoki, M. Perc, G. Szabó, Evolutionary establishment of moral and double moral standards through spatial interactions, PLoS Comput. Biol., 6 (2010), e1000758. https://doi.org/10.1371/journal.pcbi.1000758 doi: 10.1371/journal.pcbi.1000758
|
| [33] |
D. Helbing, A. Szolnoki, M. Perc, G. Szabó, Punish, but not too hard: how costly punishment spreads in the spatial public goods game, New J. Phys., 12 (2010), 083005. https://doi.org/10.1088/1367-2630/12/8/083005 doi: 10.1088/1367-2630/12/8/083005
|
| [34] |
X. Chen, A. Szolnoki, M. Perc, Probabilistic sharing solves the problem of costly punishment, New J. Phys., 16 (2014), 083016. https://doi.org/10.1088/1367-2630/16/8/083016 doi: 10.1088/1367-2630/16/8/083016
|
| [35] | L. Stella, W. Baar, D. Bauso, Lower network degrees promote cooperation in the prisoner's dilemma with environmental feedback, IEEE Contr. Syst. Lett., 6 (2022). https://doi.org/10.1109/LCSYS.2022.3175402 |
| [36] | L. Stella, D. Bauso, The Impact of irrational behaviours in the optional prisoner's dilemma with game-environment feedback, Int. J. Robust Nonlinear Control, 2022 (2022). https://doi.org/10.1002/rnc.5935 |
| [37] |
X. R. Mao, G. Marion, E. Renshaw, Environmental Brownian noise suppresses explosions in population dynamics, Stochastic Processes Appl., 97 (2002), 95–110. https://doi.org/10.1016/S0304-4149(01)00126-0 doi: 10.1016/S0304-4149(01)00126-0
|
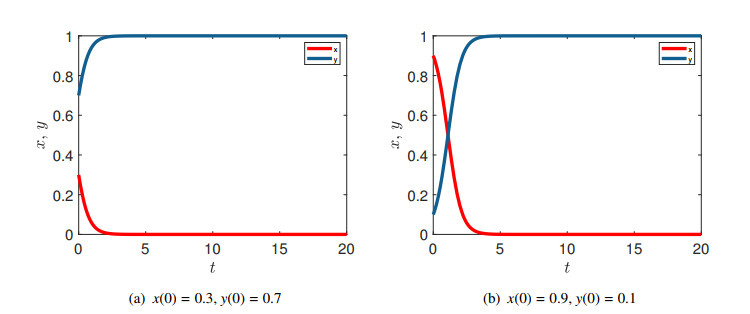














Figures(15)

Yifei Wang, Xinzhu Meng. Evolutionary game dynamics of cooperation in prisoner's dilemma with time delay[J]. Mathematical Biosciences and Engineering, 2023, 20(3): 5024-5042. doi: 10.3934/mbe.2023233







 DownLoad:
DownLoad: