Citation: Ning Li, Jining Zhou. Screening and validating the immune-related gene expression signatures in peripheral blood mononuclear cells of nonischaemic cardiomyopathy[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2330-2347. doi: 10.3934/mbe.2020124

| [1] | M. Jessup, T. H. Marwick, P. Ponikowski, A. A. Voors, C. W. Yancy, 2016 ESC and ACC/AHA/HFSA heart failure guideline update—what is new and why is it important?, Nat. Rev. Cardiol., 13 (2016), 623-628. |
| [2] | S. V. Konstantinides, G. Meyer, C. Becattini, H. Bueno, G. J. Geersing, V. P. Harjola, et al., 2019 ESC Guidelines for the diagnosis and management of acute pulmonary embolism developed in collaboration with the European Respiratory Society (ERS), Eur. Heart J., 27 (2019), 2129-2200. |
| [3] | F. Ali-Ahmed, F. Dalgaard, S. M. Al-Khatib, Sudden cardiac death in patients with myocarditis: Evaluation, risk stratification, and management, Am. Heart J., 220 (2019), 29-40. |
| [4] | C. Wojciechowska, J. Wodniecki, R. Wojnicz, E. Romuk, W. Jachec, A. Tomasik, et al., Neopterin and beta-2 microglobulin relations to immunity and inflammatory status in nonischemic dilated cardiomyopathy patients, Mediators Inflamm., 2014 (2014), 585067. |
| [5] | Y. Zhang, J. Bauersachs, H. F. Langer, Immune mechanisms in heart failure,. Eur. J. Heart Fail, 19 (2017), 1379-1389. |
| [6] | W. Chen, N. G. Frangogiannis, Fibroblasts in post-infarction inflammation and cardiac repair, Biochim. Biophys Acta, 1822 (2013), 945-953. |
| [7] | T. Shioi, A. Matsumori, Y. Kihara, M. Inoko, K. Ono, Y. Iwanaga, et al., Increased expression of interleukin-1 beta and monocyte chemotactic and activating factor/monocyte chemoattractant protein-1 in the hypertrophied and failing heart with pressure overload, Circ. Res., 18 (1997), 664-671. |
| [8] | B. Coles, C. A. Fielding, S. Rose-John, J. Scheller, S. A. Jones, V. B. O'Donnell, Classic interleukin-6 receptor signaling and interleukin-6 trans-signaling differentially control angiotensin Ⅱ-dependent hypertension, cardiac signal transducer and activator of transcription-3 activation, and vascular hypertrophy in vivo, Am. J. Pathol., 171 (2007), 315-325. |
| [9] |
M. Afanasyeva, D. Georgakopoulos, D. F. Belardi, D. Bedja, D. Fairweather, Y. Wang, et al., Impaired up-regulation of CD25 on CD4+ T cells in IFN-gamma knockout mice is associated with progression of myocarditis to heart failure, Proc. Natl. Acad. Sci. U S A, 102 (2005), 180-185. doi: 10.1073/pnas.0408241102

|
| [10] | S. D'Amore, M. Vacca, M. Cariello, G. Graziano, A. D'Orazio, R. Salvia, et al., Genes and miRNA expression signatures in peripheral blood mononuclear cells in healthy subjects and patients with metabolic syndrome after acute intake of extra virgin olive oil, Biochim. Biophys Acta, 1861, (2016), 1671-1680. |
| [11] | G. Serino, F. Sallustio, C. Curci, S. N. Cox, F. Pesce, G. De Palma, et al., Role of let-7b in the regulation of N-acetylgalactosaminyltransferase 2 in IgA nephropathy, Nephrol. Dial. Transplant, 30 (2015), 1132-1139. |
| [12] | N. Li, L. Li, Y. S. Chen, The identification of core gene expression signature in Hepatocellular Carcinoma, Oxid. Med. Cell Longev., 2018 (2018), 3478305. |
| [13] | L. Zhang, Y. Yang, L. Cheng, Y. Cheng, H. H. Zhou, Z. R. Tan, Identification of common genes refers to colorectal carcinogenesis with paired cancer and noncancer samples, Dis. Markers, 2018 (2018), 3452739. |
| [14] | P. Gaudet, N. Skunca, J. C. Hu, C. Dessimoz, Primer on the gene ontology, Methods Mol. Biol., 1446 (2017), 25-37. |
| [15] | M Kanehisa, The KEGG database, Novartis Found Symp., 247 (2002), 91-101; discussion 101-103, 119-128, 244-152. |
| [16] | D. W. Huang, B. T. Sherman, Q. Tan, J. R. Collins, W. G. Alvord, J. Roayaei, et al., The DAVID gene functional classification tool: A novel biological module-centric algorithm to functionally analyze large gene lists, Genome Biol., 8 (2007), R183. |
| [17] |
D. Szklarczyk, A. Franceschini, S. Wyder, K. Forslund, D. Heller, J. Huerta-Cepas, et al., STRING v10: Protein-protein interaction networks, integrated over the tree of life, Nucleic Acids Res., 43 (2015), D447-452. doi: 10.1093/nar/gku1003
|
| [18] | J. Zhang, M. Huang, S. Shen, X. Wu, X. Wu, P. Lv, et al., Qiliqiangxin attenuates isoproterenol-induced cardiac remodeling in mice, Am. J. Transl. Res., 9 (2017), 5585-5593. |
| [19] | S. C. Chang, S. Ren, C. D. Rau, J. J. Wan, Isoproterenol-induced heart failure mouse model using osmotic pump implantation, Methods Mol. Biol.,1816 (2018), 207-220. |
| [20] | S. Desjardins, E. Belkai, D. Crete, L. Cordonnier, J. M. Scherrmann, F. Noble, et al., Effects of chronic morphine and morphine withdrawal on gene expression in rat peripheral blood mononuclear cells, Neuropharmacology, 55 (2008), 1347-1354. |
| [21] | N. Hamdani, C. Franssen, A. Lourenco, I. Falcao-Pires, D. Fontoura, S. Leite, et al., Myocardial titin hypophosphorylation importantly contributes to heart failure with preserved ejection fraction in a rat metabolic risk model, Circ. Heart Fail, 6 (2013),1239-1249. |
| [22] | A. Kadish, A. Dyer, J. P. Daubert, R. Quigg, N. A. Estes, K. P. Anderson, et al., Prophylactic defibrillator implantation in patients with nonischemic dilated cardiomyopathy, N Engl. J. Med., 350 (2004), 2151-2158. |
| [23] | P. Richardson, W. McKenna, M. Bristow, B. Maisch, B. Mautner, J. O'Connell, et al., Report of the 1995 World Health Organization/International Society and Federation of Cardiology Task Force on the Definition and Classification of cardiomyopathies, Circulation, 93 (1996), 841-842. |
| [24] | G. Poglajen, G. Zemljic, S. Frljak, A. Cerar, V. Androcec, M. Sever, et al., Stem cell therapy in patients with chronic nonischemic heart failure, Stem Cells Int., 2018, (2018), 6487812. |
| [25] | S. Sumitomo, K. Fujio, T. Okamura, K. Yamamoto, Egr2 and Egr3 are the unique regulators for systemic autoimmunity, Jakstat, 2 (2013), e23952. |
| [26] | H. Wang, L. Hou, D. Kwak, J. Fassett, X. Xu, A. Chen, et al., Increasing regulatory T cells with Interleukin-2 and Interleukin-2 antibody complexes attenuates lung inflammation and heart failure progression, Hypertension, 68 (2016), 114-122. |
| [27] | N. Li, H. Bian, J. Zhang, X. Li, X. Ji, Y Zhang, The Th17/Treg imbalance exists in patients with heart failure with normal ejection fraction and heart failure with reduced ejection fraction, Clin. Chim. Acta, 411 (2010), 1963-1968. |
| [28] | R. M. Parkinson, S. L. Collins, M. R. Horton, J. D. Powell, Egr3 induces a Th17 response by promoting the development of gammadelta T cells, PLoS One, 9 (2014), e87265. |
| [29] | N. Glezeva, S. Horgan, J. A. Baugh, Monocyte and macrophage subsets along the continuum to heart failure: Misguided heroes or targetable villains?, J. Mol. Cell Cardiol., 89 (2015), 136-145. |
| [30] | H. R. Zandbergen, U. C. Sharma, S. Gupta, J. W. Verjans, S. van den Borne, S. Pokharel, et al., Macrophage depletion in hypertensive rats accelerates development of cardiomyopathy, J. Cardiovasc. Pharmacol. Ther., 14 (2009), 68-75. |
| [31] | R. K. Braun, J. M. Koch, T. A. Hacker, D. Pegelow, J. Kim, A. N. Raval, et al., Cardiopulmonary and histological characterization of an acute rat lung injury model demonstrating safety of mesenchymal stromal cell infusion, Cytotherapy, 18 (2016), 536-545. |
| [32] | H. Kai, F. Kuwahara, K. Tokuda, T. Imaizumi, Diastolic dysfunction in hypertensive hearts: Roles of perivascular inflammation and reactive myocardial fibrosis, Hypertens Res., 28 (2005), 483-490. |
| [33] | J. Patel, E. McNeill, G. Douglas, A. B. Hale, J. de Bono, R. Lee, et al., RGS1 regulates myeloid cell accumulation in atherosclerosis and aortic aneurysm rupture through altered chemokine signalling, Nat. Commun., 6 (2015), 6614. |
| [34] | L. Chavez-Sanchez, A. Madrid-Miller, K. Chavez-Rueda, M. V. Legorreta-Haquet, E. Tesoro-Cruz, F. Blanco-Favela, Activation of TLR2 and TLR4 by minimally modified low-density lipoprotein in human macrophages and monocytes triggers the inflammatory response, Hum. Immunol., 71 (2010), 737-744. |
| [35] | L. Gullestad, P. Aukrust, Review of trials in chronic heart failure showing broad-spectrum anti-inflammatory approaches, Am. J. Cardiol., 95 (2005), 17C-23C; discussion 38C-40C. |
| [36] | P. Alvarez, A. Briasoulis, Immune modulation in heart failure: The promise of novel biologics. |
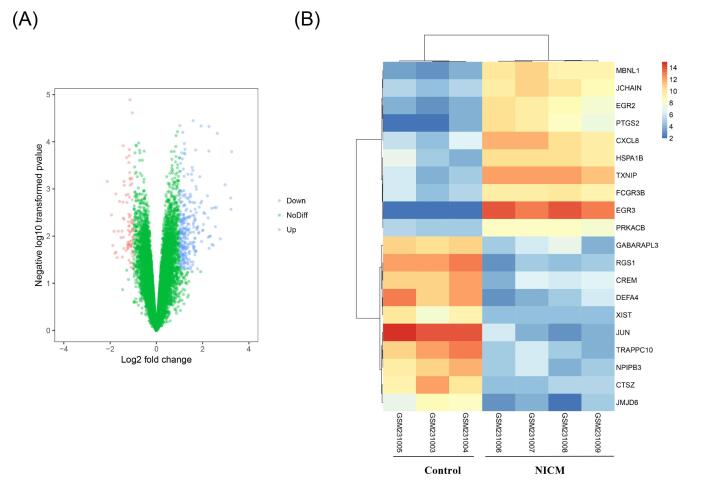
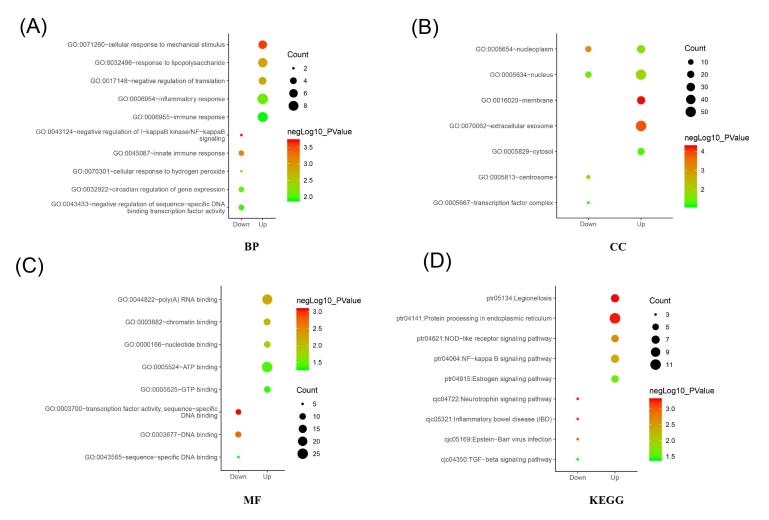
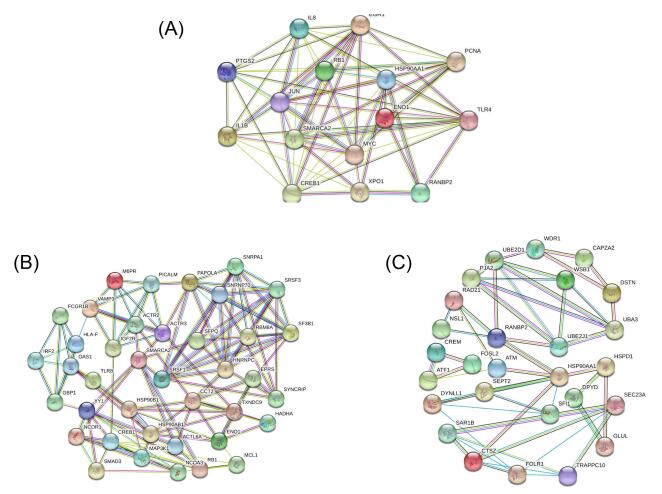
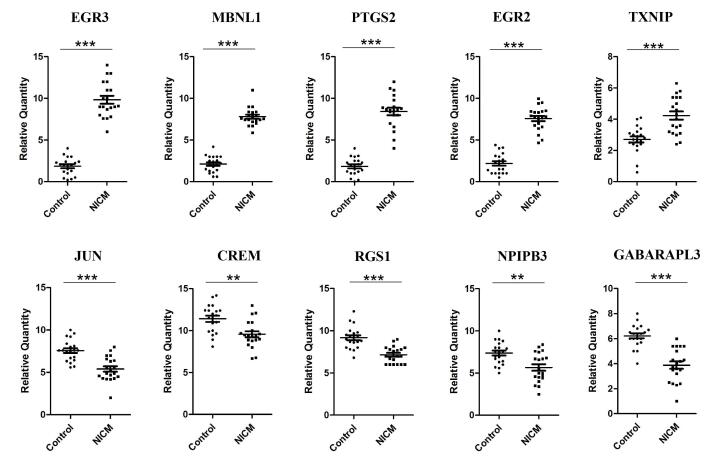
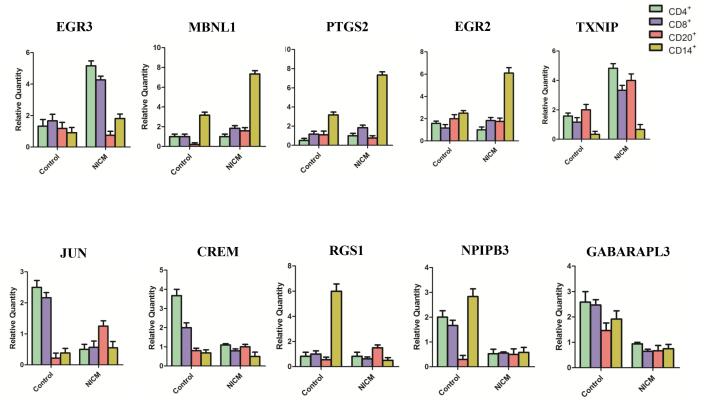
Figures(5) / Tables(4)

Ning Li, Jining Zhou. Screening and validating the immune-related gene expression signatures in peripheral blood mononuclear cells of nonischaemic cardiomyopathy[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2330-2347. doi: 10.3934/mbe.2020124







 DownLoad:
DownLoad: