Citation: Xiaochen Sheng, Weili Xiong. Soft sensor design based on phase partition ensemble of LSSVR models for nonlinear batch processes[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1901-1921. doi: 10.3934/mbe.2020100

| [1] | X. Wang, H. Liu, Soft sensor based on stacked auto-encoder deep neural network for air preheater rotor deformation prediction, Adv. Eng. Inf., 36 (2018), 112-119. |
| [2] | W. Shao, X. Tian, Adaptive soft sensor for quality prediction of chemical processes based on selective ensemble of local partial least squares models, Chem. Eng. Res. Des., 95 (2015), 113-132. |
| [3] | B. Bidar, J. Sadeghi, F, Shahraki, M. M. Khalilipour, Data-driven soft sensor approach for online quality prediction using state dependent parameter models, Chemom. Intell. Lab. Syst., 162 (2017), 130-141. |
| [4] | J. Zheng, Z. Song, Semisupervised learning for probabilistic partial least squares regression model and soft sensor application, J. Process Control, 64 (2018), 123-131. |
| [5] | D. Wang, J. Liu, R. Srinivasan, Data-Driven soft sensor approach for quality prediction in a refining process, IEEE Trans. Ind. Inf., 6 (2010), 11-17. |
| [6] | H. Kaneko, M. Arakawa, K. Funatsu, Development of a new soft sensor method using independent component analysis and partial least squares, AIChE J., 55 (2010), 87-98. |
| [7] | M. Zounemat-Kermani, D. Stephan, R. Hinkelmann, Multivariate NARX neural network in prediction gaseous emissions within the influent chamber of wastewater treatment plants, Atmos. Pollut. Res., 10 (2019), 1812-1822. |
| [8] | R. Sharmin, U. Sundararaj, S. Shah, L.V. Griend, Y. J. Sun, Inferential sensors for estimation of polymer quality parameters: Industrial application of a PLS-based soft sensor for a LDPE plant, Chem. Eng. Sci., 61 (2006), 6372-6384. |
| [9] | M. J. Willis, G. A. Montague, C. Di. Massimo, M. T. Tham, A. J. Morris, Artificial neural networks in process estimation and control, Automatica, 28 (1992), 1181-1187. |
| [10] | C. Cortes, V. Vapnik, Support Vector Network, Mach. Learn., 20 (1995), 273-297. |
| [11] | J. Tan, S. Li, Z. Zhang, C. X. Chen, W. Chen, H. Tang, et al., Identification of hormone binding proteins based on machine learning methods, Math. Biosci. Eng., 16 (2019), 2466-2480. |
| [12] | W. Xiong, W. Zhang, B. Xu, B. Huang, JITL based MWGPR soft sensor for multi-mode process with dual-updating strategy, Comput. Chem. Eng., 90 (2016), 260-267. |
| [13] | P. Facco, F. Bezzo, M. Barolo, Nearest-neighbor method for the automatic maintenance of multivariate statistical soft sensors in batch processing, Ind. Eng. Chem. Res., 49 (2010), 2336-2347. |
| [14] | V. Ferreira, F. A. A. Souza, R. Araujo, Semi-supervised soft sensor and feature ranking based on co-regularised least squares regression applied to a polymerization batch process, IEEE Int. Conf. Ind. Inf., 2017, 257-262. |
| [15] | P. Nomikos, J. F. Macgregor, Monitoring batch processes using multi-way principal component analysis, AIChE J., 40 (2010), 1361-1375. |
| [16] | Z. Lv, Q. Jiang, X. Yan, Batch process monitoring based on multisubspace multiway principal component analysis and time-series Bayesian inference, Ind. Eng. Chem. Res., 53 (2014), 6457-6466. |
| [17] | L. P. L. De Oliveira, D. Marcondes Filho, Monitoring batch processes with an incomplete set of variables, Int. J. Adv. Manuf. Technol., 94 (2018), 2515-2523. |
| [18] | C. Flavio, H. M. M. Van, Fast multiway partial least squares regression, IEEE Trans. Biomed. Eng., 66 (2019), 433-443. |
| [19] | X. Chen, X. Gao, Y. Zhang, Y. Qi, Enhanced batch process monitoring and quality prediction based on multi-phase multi-way partial least squares, IEEE International Conference on Intelligent Computing and Intelligent Systems, 2010. Available from: https://ieeexplore_ieee.xilesou.top/abstract/document/5658834. |
| [20] | H. Jin, X. Chen, L. Wang, K. Yang, L. Wu, Adaptive soft sensor development based on online ensemble Gaussian process regression for nonlinear time-varying batch processes, Ind. Eng. Chem. Res., 54 (2015), 7320-7345. |
| [21] | M. Sun, H. Yang, Gaussian process ensemble soft-sensor modeling based on improved Bagging algorithm, CIESC J., 67 (2016), 1386-1391. |
| [22] | C. Wang, X. Bai, Boosting learning algorithm for stock price forecasting, IOP Conf. Ser.: Mater. Sci. Eng., 322 (2018). |
| [23] | S. Tasnim, A. Rahman, A. M. T. Oo, M. E. Haque, Wind power prediction using cluster based ensemble regression, Int. J. Comput. Intell. Appl., 16 (2017), 1750026. |
| [24] | L. A. Gabralla, A. Abraham, Prediction of oil prices using bagging and random subspace, Proceedings of the Fifth International Conference on Innovations in Bio-Inspired Computing and Applications IBICA 2014, 2014. Available from: https://link_springer.xilesou.top/. |
| [25] | Y. Sun, Y. He, X. Ji, Soft sensor modeling of the penicillin fermentation based on FCM clustering and LS-SVM, 2010 International Conference on Computer Application & System Modeling, 2010. Available from: https://ieeexplore_ieee.xilesou.top/abstract/document/5622912. |
| [26] | L. Wang, H. Jin, X. Chen, J. Dai, K. Yang, D. Zhang, Soft Sensor Development based on the hierarchical ensemble of Gaussian process regression models for nonlinear and non-Gaussian chemical processes, Ind. Eng. Chem. Res., 55 (2016), 7704-7719. |
| [27] | L. Yao, Z. Ge, Online updating soft sensor modeling and industrial application based on selectively integrated moving window approach, IEEE Trans. Instrum. Meas., 66 (2017), 1985-1993. |
| [28] | H. Kaneko, K. Funatsu, Adaptive soft sensor based on online support vector regression and Bayesian ensemble learning for various states in chemical plants, Chemom. Intell. Lab. Syst., 137 (2014), 57-66. |
| [29] | Y. Liu, J. Chen, Integrated soft sensor using just-in-time support vector regression and probabilistic analysis for quality prediction of multi-grade processes, J. Process Control, 23 (2013), 793-804. |
| [30] | S. Xu, X. An, X. Qiao, L. Zhu, L. Li, Multi-output least-squares support vector regression machines, Pattern Recognit. Lett., 34 (2013), 1078-1084. |
| [31] | A. P. Verbyla, A note on model selection using information criteria for general linear models estimated using REML, Aust. N. Z. J. Stat., 61 (2019), 39-50. |
| [32] | N. Pillai, S. L. Schwartz, T. Ho, A. Dokoumetzidis, R. Bies, I. Freedman, Estimating parameters of nonlinear dynamic systems in pharmacology using chaos synchronization and grid search, J. Pharmacokinet. Pharmacodyn., 46 (2019), 193-210. |
| [33] | V. Vakharia, R. Gujar, Prediction of compressive strength and portland cement composition using cross-validation and feature ranking techniques, Constr. Build. Mater., 225 (2019), 292-301. |
| [34] | P. Arena; S. Baglio, L. Fortuna, G. Manganaro, Self-organization in a two-layer CNN, IEEE Trans. Circuits Syst. I Fundam. Theory Appl., 45 (1998), 157-162. |
| [35] | M. Wang, H. Chen, H. Li, Z. Cai, X. Zhao, C. Tong, et al., Grey wolf optimization evolving kernel extreme learning machine: application to bankruptcy prediction, Eng. Appl. Artif. Intell., 63 (2017), 54-68. |
| [36] | S. W. Lin, K. C. Ying, S. C. Chen, Z. J. Lee, Particle swarm optimization for parameter determination and feature selection of support vector machines, Expert Syst. Appl., 35 (2018), 1817-1824. |
| [37] | C. Wu, F. Yang, Y. Wu, R. Han, Prediction of crime tendency of high-risk personnel using C5.0 decision tree empowered by particle swarm optimization, Math. Biosci. Eng., 16 (2019), 4135-4150. |
| [38] | X. Kong, X. Che, R. Su, C. Zhang, Q. Yao, X. Shi, A new technique for rapid assessment of eutrophication status of coastal waters using a support vector machine, Chin. J. Oceanol. Limnol., 36 (2018), 1-14. |
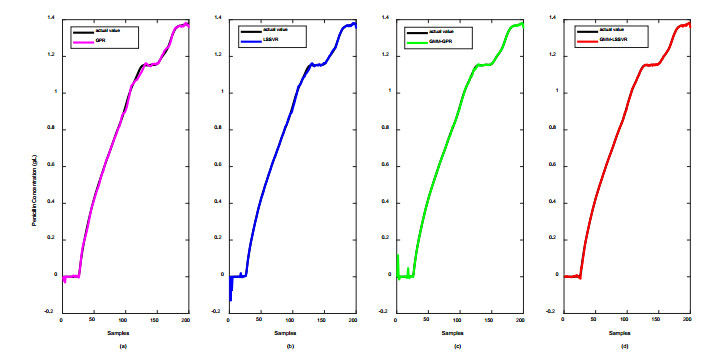
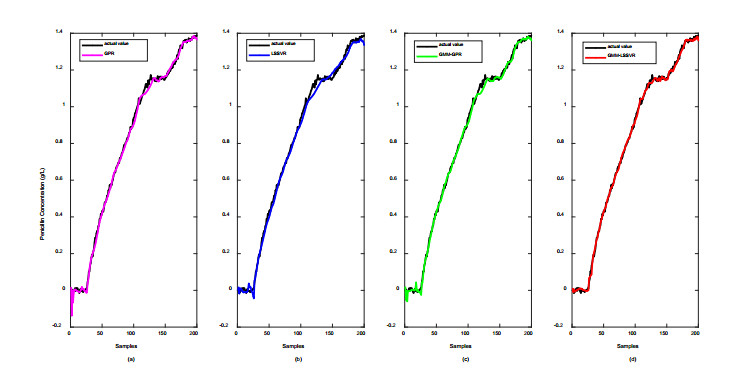
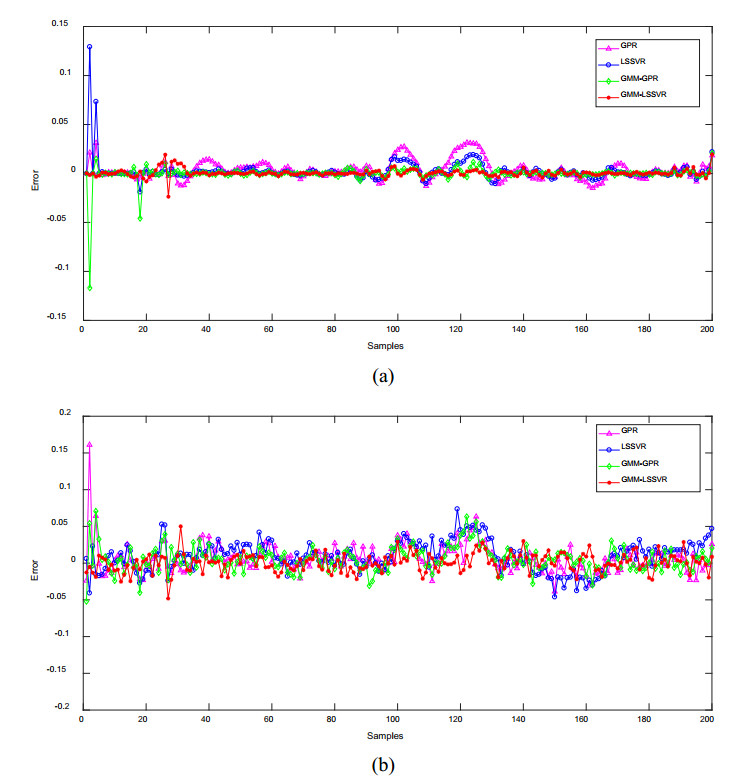
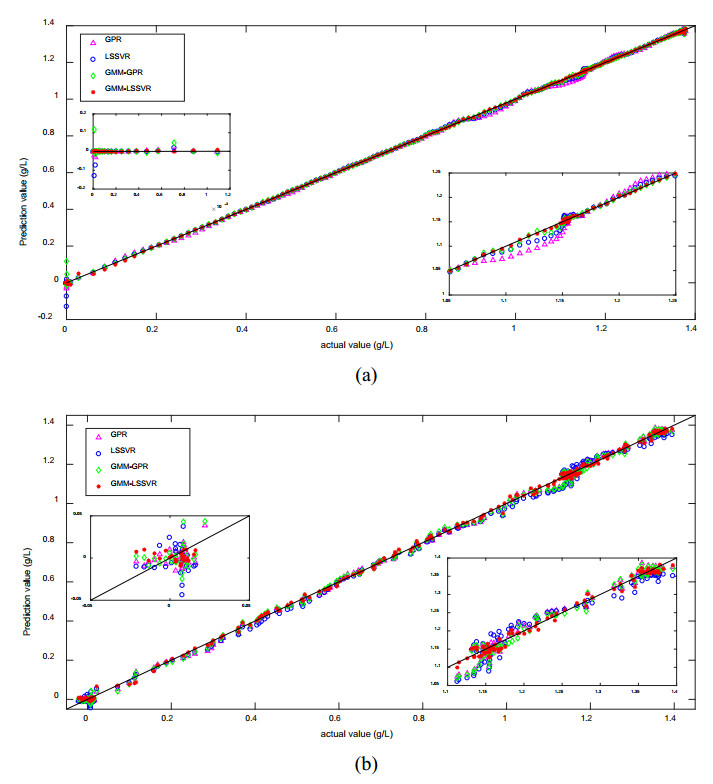
Figures(12) / Tables(2)

Xiaochen Sheng, Weili Xiong. Soft sensor design based on phase partition ensemble of LSSVR models for nonlinear batch processes[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1901-1921. doi: 10.3934/mbe.2020100







 DownLoad:
DownLoad: