Citation: Erli Yang, Xiaobei Zhang, Qiangsheng Chen, Chandong Ding. Tumor necrosis factor-related apoptosis-inducing ligand regulate the accumulation of extracelluar matrix in pulmonary artery by activating the phosphorylation of Smad2/3[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1372-1380. doi: 10.3934/mbe.2020069

| [1] | A. B. Waxman and R. T. Zamanian, Pulmonary arterial hypertension: New insights into the optimal role of current and emerging prostacyclin therapies, Am. J. Cardiol., 111 (2013), 1A-16A. |
| [2] | L. J. Rubin, Primary pulmonary hypertension, N. Engl. J. Med., 336 (1997), 111-117. |
| [3] | J. R. Runo and J. E. Loyd, Primary pulmonary hypertension, Lancet, 361 (2003), 1533-1544. |
| [4] | J. Hänze, N. Weissmann, F. Grimminger, et al., Cellular and molecular mechanisms of hypoxia inducible factor driven vascular remodeling, Thromb. Haemostasis, 97 (2007), 774-787. |
| [5] | P. M. Hassoun, Deciphering the "matrix" in pulmonary vascular remodeling, Eur. Respir. J., 25 (2005), 778-779. |
| [6] | E. Arciniegas, M. G. Frid, I. S. Douglas, et al., Perspectives on endothelial to mesenchymal transition: Potential contribution to vascular remodeling in chronic pulmonary hypertension, Am. J. Physiol. Lung Cell. Mol. Physiol., 293 (2007), L1-L8. |
| [7] |
G. Burgstaller, B. Oehrle, M. Gerckens, et al., The instructive extracellular matrix of the lung: Basic composition and alterations in chronic lung disease, Eur. Respir. J., 50 (2017), 1601805. doi: 10.1183/13993003.01805-2016

|
| [8] | V. Jurisic, T. Srdic-Rajic, G. Konjevic, et al., TNF-α induced apoptosis is accompanied with rapid CD30 and slower CD45 shedding from K-562 cells, J. Membr. Biol., 293 (2011), 115-122. |
| [9] | V. Jurisic, T. Terzic, S. Colic, et al., The concentration of TNF-α correlate with number of inflammatory cells and degree of vascularization in radicular cysts, Oral Dis., 14 (2008), 600-605. |
| [10] | A. Lawrie, A. G. Hameed, J. Chamberlain, et al., Paigen diet-fed apolipoprotein E knockout mice develop severe pulmonary hypertension in an interleukin-1-dependent manner, Am. J. Pathol., 179 (2011), 1693-1705. |
| [11] | N. M. Robertson, M. Rosemiller, R. G. Lindemeyer, et al., TRAIL in the Airways, Vitam. Horm., 67 (2004), 149-167. |
| [12] |
B. R. Gochuico, J. Zhang, B. Y. Ma, et al., TRAIL expression in vascular smooth muscle, Am. J. Physiol. Lung Cell. Mol. Physiol., 278 (2000), L1045-L1050. doi: 10.1152/ajplung.2000.278.5.L1045
|
| [13] | A. Lawrie, E. Waterman, M. Southwood, et al., Evidence of a role for osteoprotrgerin in the pathogenesis of pulmonary arterial hypertension, Am. J. Pathol., 172 (2008), 256-264. |
| [14] | X. D. Zhang, T. Nguyen, W. D. Thomas, et al., Mechanisms of resistance of normal cells to TRAIL induced apoptosis vary between different cell types, FEBS Lett., 482 (2000), 193-199. |
| [15] |
R. Di Pietro, M. A. Mariggio, S. Guarnieri, et al., Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand (TRAIL) Regulates Endothelial Nitric Oxide Synthase (eNOS) Activity and Its Localization Within the Human Vein Endothelial Cells (HUVEC) in Culture, J. Cell. Biochem., 97 (2006), 782-794. doi: 10.1002/jcb.20686
|
| [16] | H. Liu, E. Yang, X. Lu, et al., Serum levels of tumor necrosis factor-related apoptosis-inducing ligand correlate with the severity of pulmonary hypertension, Pulm. Pharmacol. Ther., 33 (2015), 39-46. |
| [17] | A. G. Hameed, N. D. Arnold, J. Chamberlain, et al., Inhibition of tumor necrosis factor-related Apoptosis-inducing ligand reverses experimental pulmonary hypertension, J. Exp. Med., 216 (2012), 1919-1935. |
| [18] | G. S. Wu, TRAIL as a target in anti-cancer therapy, Cancer Lett., 285 (2009), 1-5. |
| [19] | S. R. Wiley, K. Schooley, P. J. Smolak, et al., Identification and characterization of a new member of the TNF family that induces apoptosis, Immunity, 3 (1995), 673-682. |
| [20] | H. B. Schiller, I. E. Fernandez, G. Burgstaller, et al., Time- and compartment-resolved proteome profiling of the extracellular niche in lung injury and repair, Mol. Syst. Biol., 11 (2015), 819. |
| [21] | H. N. LeBlanc, A. Ashkenazi, Apo2L/TRAIL and its death and decoy receptors, Cell Death Differ., 10 (2003), 66-75. |
| [22] |
P. Secchiero, F. Corallini, M. G. di Iasio, et al., TRAIL counteracts the proadhesive activity of inflammatory cytokines in endothelial cells by down-modulating CCL8 and CXCL10 chemokine expression and release, Blood, 105 (2005), 3413-3419. doi: 10.1182/blood-2004-10-4111
|
| [23] | A. Almasan and A. Ashkenazi, Apo2L/TRAIL: Apoptosis signaling, biology, and potential for cancer therapy, Cytokine Growth Factor Rev., 14 (2003), 337-348. |
| [24] | P. Secchiero, A. Gonelli, E. Carnevale, et al., TRAIL promotes the survival and proliferation of primary human vascular endothelial cells by activating the Akt and Erk pathway, Circulation, 107 (2003), 2250-2256. |
| [25] | J. P. Sheridan, S. A. Marsters, R. M. Pitti, et al., Control of TRAIL-induced apoptosis by a family of signaling and decoy receptors, Science, 277 (1997), 818-821. |
| [26] | C. Falschlehner, C. H. Emmerich, B. Gerlach, et al., TRAIL signalling: Decisions between life and death, Int. J. Biochem. Cell Biol., 39 (2007), 1462-1475. |
| [27] | S. M. Mariani and P. H. Krammer, Differential regulation of TRAIL and CD95 ligand in transformed cells of the T and B lymphocyte lineage, Eur. J. Immunol., 28 (1998), 973-982. |
| [28] |
P. Secchiero, R. Candido, F. Corallini, et al., Systemic tumor necrosis factor-related apoptosis-inducing ligand delivery shows antiatherosclerotic activity in apolipoprotein E-null diabetic mice, Circulation, 114 (2006), 1522-1530. doi: 10.1161/CIRCULATIONAHA.106.643841
|
| [29] | X. Liu, V. R. Winrow, M. Horrocks, et al., Differential expression of TRAIL and its receptors relative to calcification in AAA, Biochem. Biophys. Res. Commun., 358 (2007), 18-23. |
| [30] | S. P. Cartland, S. W. Genner, A. Zahoor, et al., Comparative Evaluation of TRAIL, FGF-2 and VEGF-A-Induced Angiogenesis In Vitro and In Vivo, Int. J. Mol. Sci., 17 (2016), 2025. |
| [31] | J. R. Jackson, M. P. Seed, C. H. Kircher, et al., The codependence of angiogenesis and chronic inflammation, FASEB J., 11 (1997), 457-465. |
| [32] | A. A. Eddy and C. M. Giachelli, Renal expression of genes that promote interstitial inflammation and fibrosis in rats with protein-overload proteinuria, Kidney Int., 47 (1995), 1546-1557. |
| [33] | F. Verrecchia, and A. Mauviel, Transforming growth factor-beta signaling through the Smad pathway: Role in extracellular matrix gene expression and regulation, J. Invest. Dermatol., 118 (2002), 211-215. |
| [34] | A. Leask, D. J. Abraham, D. R. Finlay, et al., Dysregulation of transforming growth factor beta signaling in scleroderma: Overexpression of endoglin in cutaneous scleroderma fibroblasts, Arthritis Rheumatol., 46 (2002), 1857-1865. |
| [35] | K. J. Gordon, M. Dong, E. M. Chislock, et al., Loss of type III transforming growth factor beta receptor expression increases motility and invasiveness associated with epithelial to mesenchymal transition during pancreatic cancer progression, Carcinogenesis, 29 (2008), 252-262. |
| [36] | G. Sánchez-Duffhues, C. Hiepen, P. Knaus, et al., Bone morphogenetic protein signaling in bone homeostasis, Bone, 80 (2015), 43-59. |
| [37] | A. G. de Vinuesa, S. Abdelilah-Seyfried, P. Knaus, et al., BMP signaling in vascular biology and dysfunction, Cytokine Growth Factor Rev., 27 (2016), 65-79. |
| [38] | C. H. Heldin, K. Miyazono and P. Ten Dijke, TGF-beta signaling from cell membrane to nucleus through SMAD proteins, Nature, 390 (1997), 465-471. |
| [39] | K. Miyazono, K. Kusanagi and H. Inoue, Divergence and convergence of TGF-beta/BMP signaling, J. Cell. Physiol., 187 (2001), 265-276. |
| [40] | H. Yagita, K. Takeda, Y. Hayakawa, et al., TRAIL and its receptors as targets for cancer therapy, Cancer Sci., 95 (2004), 777-783.. |
| [41] | T. Kunieda, N. Nakanishi, T. Satoh, et al., Prognoses of primary pulmonary hypertension and chronic majorvessel thromboembolic pulmonary hypertension determined from cumulative survival curves, Intern. Med., 38 (1999), 543-546. |
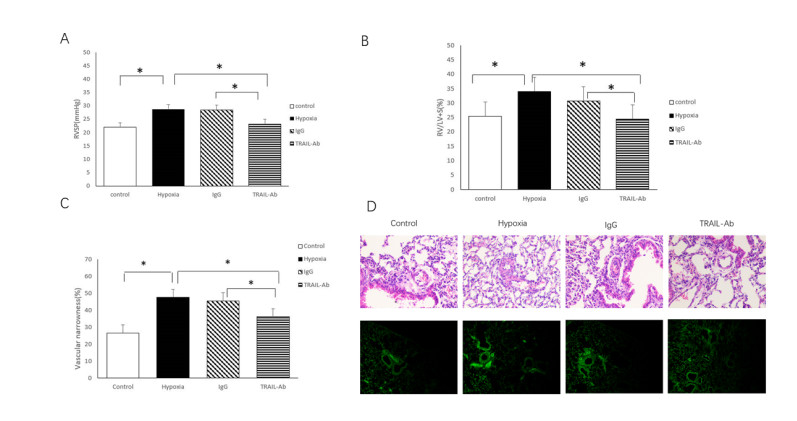
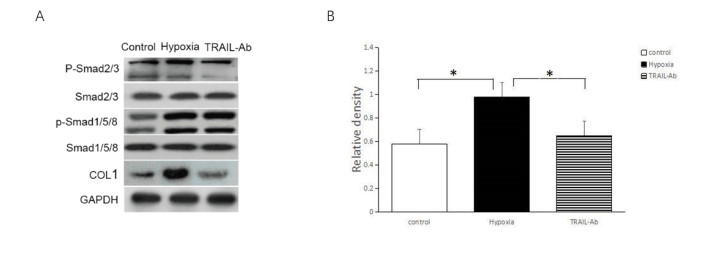
Figures(2)

Erli Yang, Xiaobei Zhang, Qiangsheng Chen, Chandong Ding. Tumor necrosis factor-related apoptosis-inducing ligand regulate the accumulation of extracelluar matrix in pulmonary artery by activating the phosphorylation of Smad2/3[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1372-1380. doi: 10.3934/mbe.2020069







 DownLoad:
DownLoad: