Citation: Hui Cao, Dongxue Yan, Ao Li. Dynamic analysis of the recurrent epidemic model[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 5972-5990. doi: 10.3934/mbe.2019299

| [1] | L. Simonsen, The global impact of influenza on morbidity and mortality, Vaccine, 17 (1999), 3–10. |
| [2] | S.W. Huang, Y.W. Hsu, D.J. Smith, et.al., Reemergence of Enterovirus 71 in 2008 in Taiwan: Dynamics of Genetic and Antigenic Evolution from 1998 to 2008, J. Clin. Microbiol., 47 (2009), 3653–3662. |
| [3] | S. Chiba, R. Kogasaka, M. Akihara, et al., Recurrent attack of rotavirus gastroenteritis after adenovirus-induced diarrhoea, Arch. Dis. Child., 54 (1979), 398–400. |
| [4] | A. Johansen, A simple model of recurrent epidemics, J. Theor. Biol., 178 (1996), 45–51. |
| [5] | B. F. Finkenstadt, A stochastic model for extinction and recurrence of epidemics: estimation and inference for measles, Biostatistics, 3(2002), 493–510. |
| [6] | A. L. Lloyd, Estimating variability in models for recurrent epidemics: assessing the use of moment closure techniques, Theor. Popul. Biol., 65 (2004), 49–65. |
| [7] | L. Stone, R. Olinky and A. Huppert, Seasonal dynamics of recurrent epidemics, Nature, 446 (2007), 533–536. |
| [8] | R. Olinky, A. Huppert and L. Stone, Seasonal dynamics and thresholds governing recurrent epidemics, J. Math. Biol., 56(2008), 827–839. |
| [9] | M. Begon, S. Telfer, M. J. Smith, et al., Seasonal host dynamics drive the timing of recurrent epidemics in a wildlife population, Proc. R. Soc. B, 276 (2009), 1063–1610. |
| [10] | J. Verdasca, M. M. Telo da Gama, A. Numes, et al., Recurrent epidemics in small world networks, J. Theor. Biol., 233 (2005), 553–561. |
| [11] | M. Zheng, C. Wang, J. Zhou, et al., Non-periodic outbreaks of recurrent epidemics and its network modelling, Sci. Rep., 5 (2015), 16010. |
| [12] | H. Liu, M. Zheng, D. Wu, et al., Hysteresis loop of nonperiodic outbreaks of recurrent epidemics, Phys. Rev., 94 (2016), 062318. |
| [13] | K. J. Engel and R. Nagel, One-Parameter Semigroups for Linear Evolution Equations, Springer, New York, 2000. |
| [14] | A. Pazy, Semigroups of Linear Operators and Applications to Partial Differential Equations, Springer, New York, 1983. |
| [15] | M. Martcheva and H. R. Thieme, Progression age enhanced backward bifurcation in an epidemic model with super-infection, J. Math. Biol., 46 (2003), 385–424. |
| [16] | H. R. Thieme, Convergence results and a Poincaré-Bendixson trichotomy for asymptotically autonomous differential equations, J. Math. Biol., 30 (1992), 755–763. |
| [17] | J. K. Hale, Theory of Function Differential Equations, Springer, Heidelberg, 1977. |
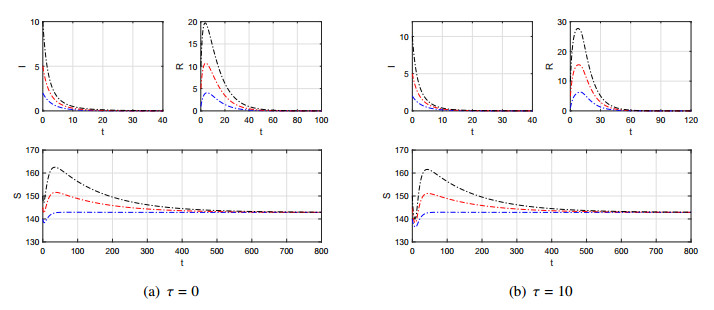
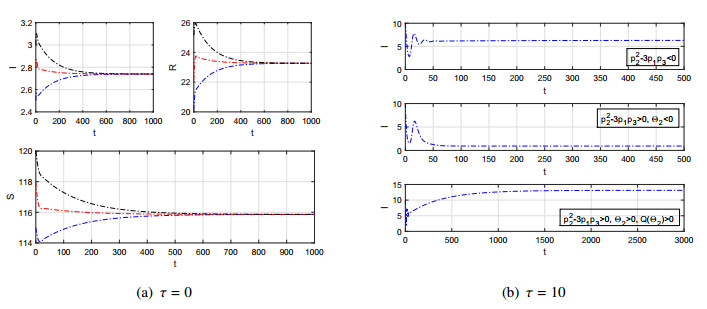
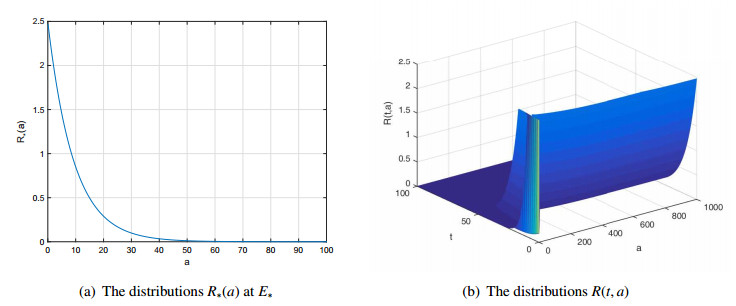
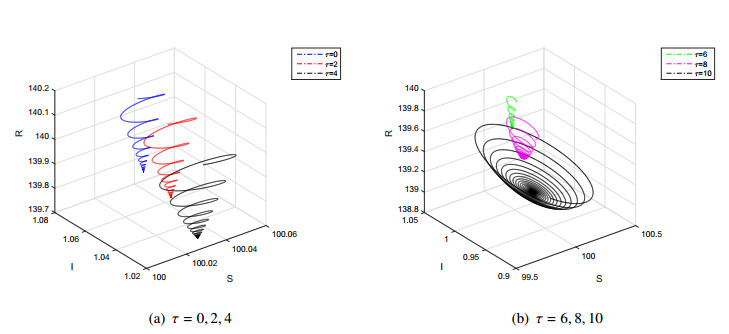
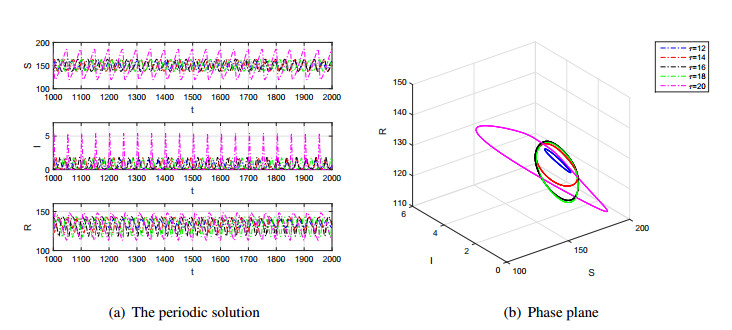
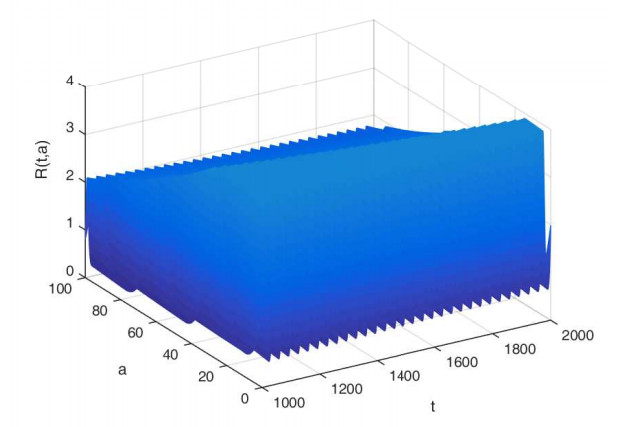
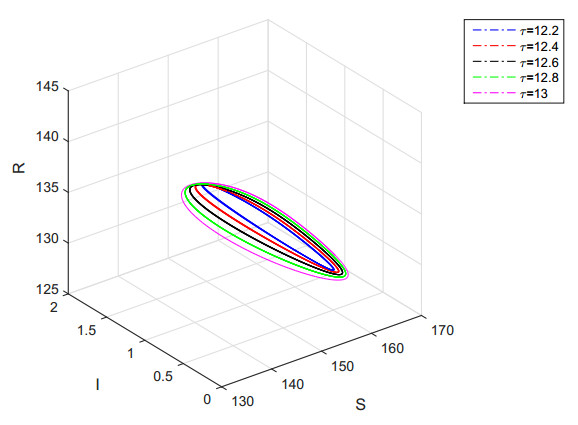
Figures(7)

Hui Cao, Dongxue Yan, Ao Li. Dynamic analysis of the recurrent epidemic model[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 5972-5990. doi: 10.3934/mbe.2019299







 DownLoad:
DownLoad: