Citation: Yu Tsubouchi, Yasuhiro Takeuchi, Shinji Nakaoka. Calculation of final size for vector-transmitted epidemic model[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2219-2232. doi: 10.3934/mbe.2019109

| [1] | E. Avila-Vales and B. Buonomo, Analysis of a mosquito-borne disease transmission model with vector stages and nonlinear forces of infection, Ric. di Matem., 64 (2015), 377. |
| [2] | J. Chen, J. C. Beier and R. S. Cantrell, et al., Modeling the importation and local transmission of vector-borne diseases in florida: The case of zika outbreak in 2016, J. Theor. Biol., 455 (2018), 342–356. |
| [3] | D. Clancy and A. B. Piunovskiy, An explicit optimal isolation policy for a deterministic epidemic model, Appl. Math. Comput., 163 (2005), 1109–1121. |
| [4] | J. E. Kim, Y. Choi and C. H. Lee, Effects of climate change on plasmodium vivax malaria transmission dynamics: A mathematical modeling approach, Appl. Math. Comput., 347 (2019), 616–630, |
| [5] | C. N. Ngonghala, S. Y. D. Valle, R. Zhao and J. Mohammed-Awel, Quantifying the impact of decay in bed-net efficacy on malaria transmission, J. Theor. Biol., 363 (2014), 247 – 261, |
| [6] | S. Pandey, S. Nanda, A. Vutha and R. Naresh, Modeling the impact of biolarvicides on malaria transmission., J. Theor. Biol., 454 (2018), 396–409. |
| [7] | S. Side and M. Md. Noorani, A sir model for spread of dengue fever disease (simulation for south sulawesi, indonesia and selangor, malaysia), W. J. Model. Sim., 9 (2013), 96–105. |
| [8] | K. P. Wijaya and D. Aldila, Learning the seasonality of disease incidences from empirical data, arXiv preprint arXiv:1710.05464. |
| [9] | L. Allen, An Introduction to Mathematical Biology, Pearson/Prentice Hall, 2007. |
| [10] | L. Allen, F. Brauer and P. van den Driessche, et al., Mathematical Epidemiology, Lecture Notes in Mathematics, Springer Berlin Heidelberg, 2008. |
| [11] | O. Diekmann, H. Heesterbeek and T. Britton, Mathematical Tools for Understanding Infectious Disease Dynamics:, Princeton University Press, 2013. |
| [12] | M. Iannelli and A. Pugliese, An introduction to mathematical population dynamics : along the trail of Volterra and Lotka, no. 79 in Collana unitext, Springer, 2014. |
| [13] | N. Nakada, M. Nagata and Y. Dong, et al., Dynamics of tumor immune escape via adaptive change, Nonl. Theor. Appl., 9 (2018), 295–304. |

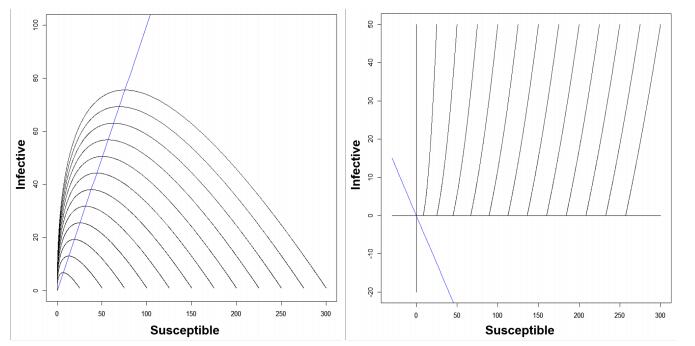
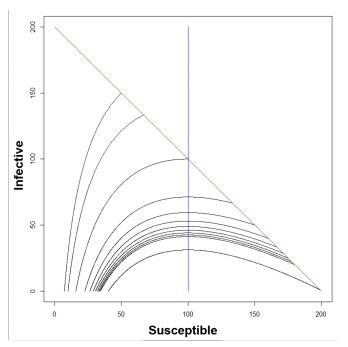
Figures(7) / Tables(1)

Yu Tsubouchi, Yasuhiro Takeuchi, Shinji Nakaoka. Calculation of final size for vector-transmitted epidemic model[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2219-2232. doi: 10.3934/mbe.2019109







 DownLoad:
DownLoad: