Citation: Weijiu Liu. A mathematical model for the robust blood glucose tracking[J]. Mathematical Biosciences and Engineering, 2019, 16(2): 759-781. doi: 10.3934/mbe.2019036

| [1] | E. Ackerman, J.W. Rosevear andW. F. McGuckin, A mathematical model of the glucose-tolerance test, Phys. Med. Biol., 9 (1964), 203–213. |
| [2] | E. Ackerman, L. C. Gatewood, J. W. Rosevear and G. D. Molnar, Model studies of blood glucose regulation, Bull. Math. Biophys., 27 (1965), 21–37. |
| [3] | B. Ashley and W. Liu, Asymptotic tracking and disturbance rejection of the blood glucose regulation system, Math. Biosci., 289 (2017), 78–88. |
| [4] | R. N. Bergman, Y. Z. Ider, C. R. Bowden and C. Cobelli, Quantitative estimation of insulin sensitivity, Am. J. Physiol. Endocrinol. Metab., 236 (1979), E667–E677. |
| [5] | R. N. Bergman, L. S. Phillips and C. Cobelli, Measurement of insulin sensitivity and β-cell glucose sensitivity from the response to intraveous glucose, J. Clin. Invest., 68 (1981), 1456–1467. |
| [6] | R. N. Bergman, D. T. Finegood and M. Ader, Assessment of insulin sensitivity in vivo, Endocrine Reviews, 6 (1985), 45–86. |
| [7] | R. N. Bergman, Toward physiological understanding of glucose tolerance, Minimal-model approach, Diabetes, 38 (1989), 1512–1527. |
| [8] | A. Bertoldo, R. R. Pencek, K. Azuma, J. C. Price, C. Kelley, C. Cobelli and D. E. Kelley, Interactions between delivery, transport, and phosphorylation of glucose in governing uptake into human skeletal muscle, Diabetes, 55 (2006), 3028–3037. |
| [9] | J. Carr, Applications of Center Manifold Theory, Applied Mathematical Sciences 35, Springer, New York, 1981. |
| [10] | C. Cobelli, G. Federspil, G. Pacini, A. Salvan and C. Scandellari, An integrated mathematical model of the dynamics of blood glucose and its hormonal control, Math. Biosci., 58 (1982), 27– 60. |
| [11] | K. Fessel, J. B. Gaither, J. K. Bower, G. Gaillard and K. Osei, Mathematical analysis of a model for glucose regulation, Mathematical Biociences and Engineering, 13(2016), 83–90. |
| [12] | L. B. Freidovich and H. K. Khalil, Performance recovery of feedback-linearization-based designs, IEEE Trans. Automat. Contr., 53 (2008), 2324–2334. |
| [13] | W. T. Garvey, L. Maianu, J. H. Zhu, G. Brechtel-Hook, P. Wallace and A. D. Baron, Evidence for defects in the tra cking and translocation of GLUT4 glucose transporters in skeltal muscle as a cause of human insulin resistance, J. Clin. Invest., 101 (1998), 2377–2386. |
| [14] | C. J. Goodner, B. C. Walike, D. J. Koerker, J. W. Ensinck, A. C. Brown, E. W. Chideckel, J. Palmer and L. Kalnasy, Insulin, glucagon, and glucose exhibit synchronous, sustained oscillations in fasting monkeys, Science, 195 (1977), 177–179. |
| [15] | O. I. Hagren and A. Tengholm, Glucose and insulin synergistically activate phosphatidylinositol 3- kinase to trigger oscillations of phosphatidylinositol 3,4,5-trisphosphate in β-cells, J. Biol. Chem., 281 (2006), 39121–39127. |
| [16] | B. C. Hansen, K. C. Jen, S. B. Pek and R. A.Wolfe, Rapid oscillations in plasma insulin, glucagon, and glucose in obese and normal weight humans. J. Clin. Endocr. Metab., 54 (1982), 785–792. |
| [17] | A. Klip and M. Vranic, Muscle, liver, and pancreas: Three Musketeers fighting to control glycemia, Am. J. Physiol. Endocrinol. Metab., 291 (2006), E1141–E1143. |
| [18] | R. Hovorka, Continuous glucose monitoring and closed-loop systems, Diabetic Med., 23 (2006), 1–12. |
| [19] | J. Huang, Nonlinear output regulation, theory and applications, Society for Industrial and Applied Mathematics, Philadelphia, 2004. |
| [20] | H. Kang, K. Han and M. Choi, Mathematical model for glucose regulation in the whole-body system, Islets, 4 (2012), 84–93. |
| [21] | D. A. Lang, D. R. Matthews, J. Peto, and R. C. Turner, Cyclic oscillations of basal plasma glucose and insulin concentrations in human beings, New Engl. J. Med., 301 (1979), 1023–1027. |
| [22] | J. Lee, R. Mukherjee and H.K. Khalil, Output feedback performance recovery in the presence of uncertainties, Syst. Control Lett., 90 (2016), 31–37. |
| [23] | J. Li, Y. Kuang and C. C. Mason, Modeling the glucose-insulin regulatory system and ultradian insulin secretory oscillations with two explicit time delays, J. Theor. Biol., 242 (2006), 722–735. |
| [24] | W. Liu and F. Tang, Modeling a simplified regulatory system of blood glucose at molecular levels, J. Theor. Biol., 252 (2008), 608–620. |
| [25] | W. Liu, C. Hsin, and F. Tang, A molecular mathematical model of glucose mobilization and uptake, Math. Biosciences, 221 (2009), 121–129. |
| [26] | W. Liu, Elementary Feedback Stabilization of the Linear Reaction Diffusion Equation and the Wave Equation, Mathematiques et Applications, Vol. 66, Springer, 2010. |
| [27] | W. Liu, Introduction to Modeling Biological Cellular Control Systems, Modeling, Simulation and Applications, Vol. 6, Springer, 2012. |
| [28] | K. Ma, H. K. Khalil and Y. Yao, Guidance law implementation with performance recovery using an extended high-gain observer, Aerosp. Sci. Technol., 24 (2013), 177–186. |
| [29] | C. D. Man, A. Caumo, R. Basu, R. A. Rizza, G. Toffolo and C. Cobelli, Minimal model estimation of glucose absorption and insulin sensitivity from oral test: validation with a tracer method, Am. J. Physiol. Endocrinol. Metab., 287 (2004), E637–E643. |
| [30] | C. D. Man, M. Campioni, K. S. Polonsky, R. Basu, R. A. Rizza, G. Toffolo and C. Cobelli, Twohour seven-sample oral glucose tolerance test and meal protocol: minimal model assessment of β-cell responsivity and insulin sensitivity in nondiabetic individuals, Diabetes, 54 (2005), 3265– 3273. |
| [31] | C. D. Man, R. A. Rizza and C. Cobelli, Meal simulation model of the glucose-insulin system, IEEE Trans. Biomed. Eng., 54 (2007), 1740–1749. |
| [32] | H. Nishimura, F. Pallardo, G. A. Seidner, S. Vannucci, I. A. Simpson and M. J. Birnbaum, Kinetics of GLUT1 and GLUT4 glucos transporters expressed in Xenopus oocytes, J. Biol. Chem., 268 (1993), 8514–8520. |
| [33] | A. E. Panteleon, M. Loutseiko, G. M. Steil and K. Rebrin, Evaluation of the effect of gain on the meal response of an automated closed-loop insulin delivery system, Diabetes, 55 (2006), 1995– 2000. |
| [34] | A. R. Sedaghat, A. Sherman and M. J. Quon, A mathematical model of metabolic insulin signaling pathways, Am. J. Physiol. Endocrinol. Metab., 283 (2002), E1084–E1101. |
| [35] | E. T. Shapiro, H. Tillil, K. S. Polonsky, V. S. Fang, A. H. Rubenstein and E. V. Cauter, Oscillations in insulin secretion during constant glucose infusion in normal man: relationship to changes in plasma glucose, J. Clin. Endocr. Metab., 67 (1988), 307–314. |
| [36] | C. Simon, G. Brandenberger and M. Follenius, Ultradian oscillations of plasma glucose, insulin, and C-peptide in man during continuous enteral nutrition, J. Clin. Endocr. Metab., 64 (1987), 669–674. |
| [37] | J. T. Sorensen, A Physiological Model of Glucose Metabolism in Man and its Use to Design and Assess Improved Insulin Therapies for Diabetes, PhD Thesis, Massachusetts Institute of Technology, 1985. |
| [38] | G. M. Steil, K. Rebrin, C. Darwin, F. Hariri and M. F. Saad, Feasibility of automating insulin delivery for the treatment of type 1 diabetes, Diabetes, 55 (2006), 3344–3350. |
| [39] | J. Sturis, E. V. Cauter, J. D. Blackman and K. S. Polonsky, Entrainment of pulsatile insulin secretion by oscillatory glucose infusion, J. Clin. Invest., 87 (1991), 439-445. |
| [40] | J. Sturis, K. S. Polonsky, E. Mosekilde and E. V. Cauter, Computer model for mechanisms underlying ultradian oscillations of insulin and glucose, Am. J. Physiol. Endocrinol. Metab., 260 (1991), E801–E809. |
| [41] | G. Toffolo and C. Cobelli, The hot IVGTT two-compartment minimal model: an improved version, Am. J. Physiol. Endocrinol. Metab., 284 (2003), E317–E321. |
| [42] | G. Toffolo, M. Campioni, R. Basu, R. A. Rizza and C. Cobelli, A minimal model of insulin secretion and kinetics to assess hepatic insulin extraction, Am. J. Physiol. Endocrinol. Metab. 290 (2006), E169–E176. |
| [43] | I. M. Tolic, E. Mosekilde and J. Sturis, Modeling the insulin-glucose feedback system: the significance of pulsatile insulin secretion, J. Theor. Biol., 207 (2000), 361–375. |
| [44] | R. C. Turner, R. R. Holman, D. Matthews, T. D. Hockaday and J. Peto, Insulin deficiency and insulin resistance interaction in diabetes: estimation of their relative contribution by feedback analysis from basal plasma insulin and glucose concentrations, Metabolism, 28 (1979), 1086– 1096. |
| [45] | O. Vahidi, K. E. Kwok, R. B. Gopaluni and L. Sun, Developing a physiological model for type II diabetes mellitus, Biochem. Eng. J., 55 (2011), 7-16. |
| [46] | O. Vahidi1, K. E. Kwok, R. B. Gopaluni and F. K. Knop, A comprehensive compartmental model of blood glucose regulation for healthy and type 2 diabetic subjects, Med. Biol. Eng. Comput., 54 (2016), 1383-1398. |
| [47] | R. R. Wolfe, J. R. Allsop and J. F. Burke, Glucose metabolism in man: Responses to intravenous glucose infusion, Metabolism, 28 (1979), 210–220. |
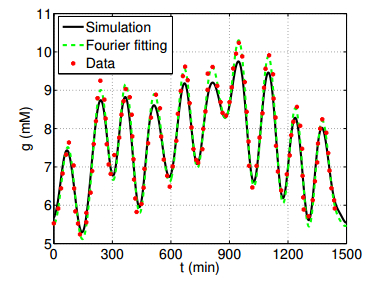
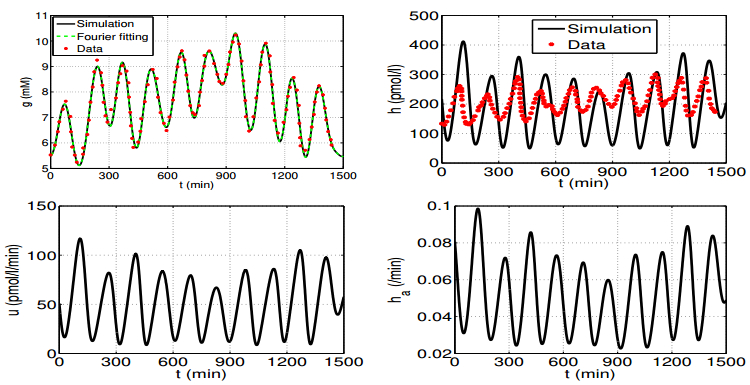
Figures(6) / Tables(1)

Weijiu Liu. A mathematical model for the robust blood glucose tracking[J]. Mathematical Biosciences and Engineering, 2019, 16(2): 759-781. doi: 10.3934/mbe.2019036







 DownLoad:
DownLoad: