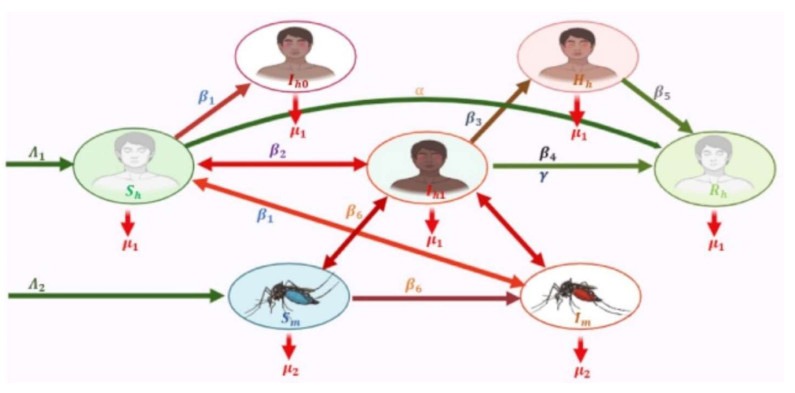
Dengue impacts 129 nations, threatens over 50% of the global population, and results in around 400 million illnesses annually. The purpose of this paper was to build the global stability and sensitivity analysis of a vector-host dengue mathematical model with compartments of symptomatic and hospitalized infected humans. Additionally, it aimed to assess the impact of the immunological response of vulnerable individuals, through the ingestion of natural foods, on the transmission of the disease. The solution's positivity and boundedness proved the model's mathematical well-posedness. To examine endemicity, the reproduction number was calculated using the next-generation technique. The Lyapunov function approach was employed to illustrate the model's global stability. Our mathematical discoveries were illustrated through numerical simulations of the dengue epidemic. The dynamical system sensitivity analysis suggests that the best way to control illness is to increase the immune system rate of susceptible hosts by consuming natural foods.
Citation: Turki D. Alharbi, Md Rifat Hasan. Global stability and sensitivity analysis of vector-host dengue mathematical model[J]. AIMS Mathematics, 2024, 9(11): 32797-32818. doi: 10.3934/math.20241569
Dengue impacts 129 nations, threatens over 50% of the global population, and results in around 400 million illnesses annually. The purpose of this paper was to build the global stability and sensitivity analysis of a vector-host dengue mathematical model with compartments of symptomatic and hospitalized infected humans. Additionally, it aimed to assess the impact of the immunological response of vulnerable individuals, through the ingestion of natural foods, on the transmission of the disease. The solution's positivity and boundedness proved the model's mathematical well-posedness. To examine endemicity, the reproduction number was calculated using the next-generation technique. The Lyapunov function approach was employed to illustrate the model's global stability. Our mathematical discoveries were illustrated through numerical simulations of the dengue epidemic. The dynamical system sensitivity analysis suggests that the best way to control illness is to increase the immune system rate of susceptible hosts by consuming natural foods.

| [1] |
O. C. Akinsulie, I. Idris, Global re-emergence of dengue fever: The need for a rapid response and surveillance, The Microbe, 4 (2024), 100107. https://doi.org/10.1016/j.microb.2024.100107 doi: 10.1016/j.microb.2024.100107

|
| [2] |
R. K. Mohapatra, P. Bhattacharjee, D. N. Desai, V. Kandi, A. K. Sarangi, S. Mishra, et al., Global health concern on the rising dengue and chikungunya cases in the American regions: countermeasures and preparedness, Health Sci. Rep., 7 (2024), e1831. https://doi.org/10.1002/hsr2.1831 doi: 10.1002/hsr2.1831
|
| [3] |
L. AL-Eitan, M. Alnemri, H. Ali, M. Alkhawaldeh, A. Mihyar, Mosquito-borne diseases: Assessing risk and strategies to control their spread in the Middle East, J Biosaf. Biosecur., 6 (2024), 1–12. https://doi.org/10.1016/j.jobb.2023.12.003 doi: 10.1016/j.jobb.2023.12.003
|
| [4] |
J. Intirach, C. Shu, X. Lv, S. Gao, N. Sutthanont, T. Chen, et al., Human parasitic infections of the class Adenophorea: global epidemiology, pathogenesis, prevention and control, Infect. Dis. Poverty, 13 (2024), 48. https://doi.org/10.1186/s40249-024-01216-1 doi: 10.1186/s40249-024-01216-1
|
| [5] |
K. Hushmandi, S. H. Saadat, S. Bokaie, H. Akbarein, S. Daneshi, M. Najafi, et al., An overview of dengue fever, Arch. Adv. Biosci., 15 (2024), 1–24. https://doi.org/10.22037/aab.v15i1.44108 doi: 10.22037/aab.v15i1.44108
|
| [6] | S. Kumar, D. Kaushik, S. K. Sharma, Autoimmune disorders: Types, symptoms, and risk factors, In: Artificial intelligence and autoimmune diseases, Singapore: Springer, 2024. https://doi.org/10.1007/978-981-99-9029-0_1 |
| [7] | D. Vilar-Compte, P. Cornejo-Juárez, E. S. Rojas, Chapter 7-Fever and rash, A Rational Approach to Clinical Infectious Diseases, 2022, 1–26. https://doi.org/10.1016/b978-0-323-69578-7.00007-7 |
| [8] |
J. Lourenço, W. Tennant, N. R. Faria, A. Walker, S. Gupta, M. Recker, Challenges in dengue research: A computational perspective, Evol. Appl., 11 (2018), 516–533. https://doi.org/10.1111/eva.12554 doi: 10.1111/eva.12554
|
| [9] |
P. Prasad, S. K. Gupta, K. K. Mahto, G. Kumar, A. Rani, I. Velan, et al., Influence of climatic factors on the life stages of Aedes mosquitoes and vectorial transmission: A review, J. Vector Borne Dis., 61 (2024), 158–166. https://doi.org/10.4103/jvbd.jvbd_42_24 doi: 10.4103/jvbd.jvbd_42_24
|
| [10] |
M. J. Pajor, B. Long, S. Y. Liang, Dengue: A focused review for the emergency clinician, Am. J. Emerg. Med., 82 (2024), 82–87. https://doi.org/10.1016/j.ajem.2024.05.022 doi: 10.1016/j.ajem.2024.05.022
|
| [11] |
C. Naing, W. Y. Ren, C. Y. Man, K. P. Fern, Q. Chua, C. N. Ning, et al., Awareness of dengue and practice of dengue control among the semi-urban community: A cross sectional survey, J. Community Health, 36 (2011), 1044–1049. https://doi.org/10.1007/s10900-011-9407-1 doi: 10.1007/s10900-011-9407-1
|
| [12] |
C. I. Siettos, L. Russo, Mathematical modeling of infectious disease dynamics, Virulence, 4 (2013), 295–306. https://doi.org/10.4161/viru.24041 doi: 10.4161/viru.24041
|
| [13] |
M. R. Hasan, A. A. Alsaiari, B. Z. Fakhurji, M. H. R. Molla, A. H. Asseri, M. A. A. Sumon, et al., Application of mathematical modeling and computational tools in the modern drug design and development process, Molecules, 27 (2022), 4169. https://doi.org/10.3390/molecules27134169 doi: 10.3390/molecules27134169
|
| [14] |
A. S. Mata, S. M. P. Dourado, Mathematical modeling applied to epidemics: an overview, Sao Paulo J. Math. Sci., 15 (2021), 1025–1044. https://doi.org/10.1007/s40863-021-00268-7 doi: 10.1007/s40863-021-00268-7
|
| [15] |
S. A. Carvalho, S. O. da Silva, I. da C. Charret, Mathematical modeling of dengue epidemic: control methods and vaccination strategies, Theory Biosci., 138 (2019), 223–239. https://doi.org/10.1007/s12064-019-00273-7 doi: 10.1007/s12064-019-00273-7
|
| [16] | M. R. Hasan, A. Hobiny, A. Alshehri, Dynamic vector-host dengue epidemic model with vector control and sensitivity analysis, Adv. Dyn. Syst. Appl., 18 (2023), 1–21. |
| [17] |
R. Rao, Z. Lin, X. Ai, J. Wu, Synchronization of epidemic systems with Neumann boundary value under delayed impulse, Mathematics, 10 (2022), 2064. https://doi.org/10.3390/math10122064 doi: 10.3390/math10122064
|
| [18] |
Q. Yang, X. Wang, X. Cheng, B. Du, Y. Zhao, Positive periodic solution for neutral-type integral differential equation arising in epidemic model, Mathematics, 11 (2023), 2701. https://doi.org/10.3390/math11122701 doi: 10.3390/math11122701
|
| [19] |
Y. Zhao, H. Lin, X. Qiao, Y. Zhao, H. Lin, X. Qiao, Persistence, extinction and practical exponential stability of impulsive stochastic competition models with varying delays, AIMS Math., 8 (2023), 22643–22661. https://doi.org/10.3934/math.20231152 doi: 10.3934/math.20231152
|
| [20] |
H. Xiao, Z. Li, Y. Zhang, H. Lin, Y. Zhao, A dual rumor spreading model with consideration of fans versus ordinary people, Mathematics, 11 (2023), 2958. https://doi.org/10.3390/math11132958 doi: 10.3390/math11132958
|
| [21] |
K. Hattaf, A new mixed fractional derivative with applications in computational biology, Computation, 12 (2024), 7. https://doi.org/10.3390/computation12010007 doi: 10.3390/computation12010007
|
| [22] |
K. Hattaf, A new class of generalized fractal and fractal-fractional derivatives with non-singular kernels, Fractal Fract., 7 (2023), 395. https://doi.org/10.3390/fractalfract7050395 doi: 10.3390/fractalfract7050395
|
| [23] |
B. Z. Naaly, T. Marijani, A. Isdory, J. Z. Ndendya, Mathematical modeling of the effects of vector control, treatment and mass awareness on the transmission dynamics of dengue fever, Comput. Method. Prog. Bio. Update, 6 (2024), 100159. https://doi.org/10.1016/j.cmpbup.2024.100159 doi: 10.1016/j.cmpbup.2024.100159
|
| [24] | A. Jana, S. K. Roy, M. H. A. Biswas, Transmission dynamics of dengue disease, awareness and control strategies, Int. J. Model. Simul., 2024. https://doi.org/10.1080/02286203.2024.2334979 |
| [25] |
D. Aldila, M. Z. Ndii, N. Anggriani, Windarto, H. Tasman, B. D. Handari, Impact of social awareness, case detection, and hospital capacity on dengue eradication in Jakarta: A mathematical model approach, Alex. Eng. J., 64 (2023), 691–707. https://doi.org/10.1016/j.aej.2022.11.032 doi: 10.1016/j.aej.2022.11.032
|
| [26] |
J. Li, H. Wan, M. Sun, Modeling the impact of awareness programs on the transmission dynamics of dengue and optimal control, Int. J. Biomath., 16 (2023), 2250072. https://doi.org/10.1142/s1793524522500723 doi: 10.1142/s1793524522500723
|
| [27] |
E. Bonyah, Y. Yuan, S. Mangal, Fractional stochastic modelling of dengue fever: The social awareness perspective, Sci. Afr., 22 (2023), e01966. https://doi.org/10.1016/j.sciaf.2023.e01966 doi: 10.1016/j.sciaf.2023.e01966
|
| [28] |
D. Aldila, C. Aulia Puspadani, R. Rusin, Mathematical analysis of the impact of community ignorance on the population dynamics of dengue, Fron.t Appl. Math. Stat., 9 (2023), 1094971. https://doi.org/10.3389/fams.2023.1094971 doi: 10.3389/fams.2023.1094971
|
| [29] |
S. K. Sood, V. Sood, I. Mahajan, Sahil, An intelligent healthcare system for predicting and preventing dengue virus infection, Computing, 105 (2023), 617–655. https://doi.org/10.1007/s00607-020-00877-8/figures/9 doi: 10.1007/s00607-020-00877-8/figures/9
|
| [30] | J. M. R. Macalalag, T. R. Y. Teng, E. P. de Lara-Tuprio, Global stability analysis of dengue model with awareness, In: Towards intelligent systems modeling and simulation, Cham: Springer, 2022. https://doi.org/10.1007/978-3-030-79606-8_14 |
| [31] |
S. Saha, G. Samanta, Analysis of a host-vector dynamics of a dengue disease model with optimal vector control strategy, Math. Comput. Simul., 195 (2022), 31–55. https://doi.org/10.1016/j.matcom.2021.12.021 doi: 10.1016/j.matcom.2021.12.021
|
| [32] |
T. K. Martheswaran, H. Hamdi, A. Al-Barty, A. A. Zaid, B. Das, Prediction of dengue fever outbreaks using climate variability and Markov chain Monte Carlo techniques in a stochastic susceptible-infected-removed model, Sci. Rep., 12 (2022, 5459. https://doi.org/10.1038/s41598-022-09489-y doi: 10.1038/s41598-022-09489-y
|
| [33] |
G. Bhuju, G. R. Phaijoo, D. B. Gurung, Sensitivity and bifurcation analysis of fuzzy SEIR-SEI dengue disease model, J. Math., 2022 (2022), 1927434. https://doi.org/10.1155/2022/1927434 doi: 10.1155/2022/1927434
|
| [34] |
Harshit, P. Harjule, Mathematical modelling and analysis of dengue transmission dynamics, Procedia Comput. Sci., 235 (2024), 539–548. https://doi.org/10.1016/j.procs.2024.04.053 doi: 10.1016/j.procs.2024.04.053
|
| [35] | S. Vaidya, Dr. A. Saraf, An optimal control with sensitivity analysis of dengue endemic using modified saturated incidence rate. https://doi.org/10.2139/ssrn.4664459 |
| [36] |
F. Guo, X. Tian, Stability analysis and simulation of a delayed dengue transmission model with logistic growth and nonlinear incidence rate, Int. J. Bifurcat. Chaos, 34 (2024), 2450028. https://doi.org/10.1142/s0218127424500287 doi: 10.1142/s0218127424500287
|
| [37] |
A. S. Leandro, W. A. Chiba de Castro, M. V. Garey, R. Maciel-de-Freitas, Spatial analysis of dengue transmission in an endemic city in Brazil reveals high spatial structuring on local dengue transmission dynamics, Sci. Rep., 14 (2024), 8930. https://doi.org/10.1038/s41598-024-59537-y doi: 10.1038/s41598-024-59537-y
|
| [38] |
Md. R. Hasan, A. Hobiny, A. Alshehri, Sensitivity analysis of vector-host dynamic dengue epidemic model, Commun. Math. Appl., 14 (2023), 1001–1017. https://doi.org/10.26713/cma.v14i2.2119 doi: 10.26713/cma.v14i2.2119
|
| [39] |
A. Abidemi, J. Ackora-Prah, H.O. Fatoyinbo, J. K. K. Asamoah, Lyapunov stability analysis and optimization measures for a dengue disease transmission model, Phy. A, 602 (2022), 127646. https://doi.org/10.1016/j.physa.2022.127646 doi: 10.1016/j.physa.2022.127646
|
| [40] |
M. A. Khan, Fatmawati, Dengue infection modeling and its optimal control analysis in East Java, Indonesia, Heliyon, 7 (2021), e06023. https://doi.org/10.1016/j.heliyon.2021.e06023 doi: 10.1016/j.heliyon.2021.e06023
|
| [41] |
J. Nawawi, D. Aldila, Mathematical assessment on the effect of hospitalization in dengue intervention, AIP Conf. Proc., 2242 (2020), 030002. https://doi.org/10.1063/5.0012142 doi: 10.1063/5.0012142
|
| [42] |
N. 'Izzati Hamdan, A. Kilicman, The development of a deterministic dengue epidemic model with the influence of temperature: A case study in Malaysia, Appl. Math. Model., 90 (2021), 547–567. https://doi.org/10.1016/j.apm.2020.08.069 doi: 10.1016/j.apm.2020.08.069
|
| [43] |
A. Abidemi, O. J. Peter, Host-vector dynamics of dengue with asymptomatic, isolation and vigilant compartments: insights from modelling, Eur. Phys. J. Plus., 138 (2023), 199. https://doi.org/10.1140/epjp/s13360-023-03823-7 doi: 10.1140/epjp/s13360-023-03823-7
|
| [44] |
R. Jan, M. A. Khan, J. F. Gómez-Aguilar, Asymptomatic carriers in transmission dynamics of dengue with control interventions, Optim. Contr. Appl. Met., 41 (2020), 430–447. https://doi.org/10.1002/oca.2551 doi: 10.1002/oca.2551
|
| [45] |
S. A. Jose, R. Raja, B. I. Omede, R. P. Agarwal, J. Alzabut, J. Cao, et al., Mathematical modeling on co-infection: transmission dynamics of Zika virus and Dengue fever, Nonlinear Dyn., 111 (2023), 4879–4914. https://doi.org/10.1007/s11071-022-08063-5 doi: 10.1007/s11071-022-08063-5
|
| [46] |
R. P. Kumar, G. S. Mahapatra, P. K. Santra, Dynamical analysis of SARS-CoV-2-Dengue co-infection mathematical model with optimum control and sensitivity analyses, Nonlinear Anal. Real, 80 (2024), 104175. https://doi.org/10.1016/j.nonrwa.2024.104175 doi: 10.1016/j.nonrwa.2024.104175
|
| [47] |
H. R. Pandey, G. R. Phaijoo, D. B. Gurung, Dengue dynamics in Nepal: A Caputo fractional model with optimal control strategies, Heliyon, 10 (2024), e33822. https://doi.org/10.1016/j.heliyon.2024.e33822 doi: 10.1016/j.heliyon.2024.e33822
|
| [48] |
M. Asaduzzaman, A. Kilicman, Abdulla-Al-Mamun, M. D. Hossain, Analysis of a novel conformable fractional order ASIR dengue transmission model in the perspective of Bangladesh, Math. Models Comput. Simul., 16 (2024), 431–456. https://doi.org/10.1134/S2070048224700157 doi: 10.1134/S2070048224700157
|
| [49] |
A. Dwivedi, R. Keval, Analysis for transmission of dengue disease with different class of human population, Epidemiol. Method., 10 (2021), 20200046. https://doi.org/10.1515/em-2020-0046 doi: 10.1515/em-2020-0046
|
| [50] |
L. Pimpi, S. W. Indratno, J. W. Puspita, E. Cahyono, Stochastic and deterministic dynamic model of dengue transmission based on dengue incidence data and climate factors in Bandung city, Commun. Biomath. Sci., 5 (2022), 78–89. https://doi.org/10.5614/cbms.2022.5.1.5 doi: 10.5614/cbms.2022.5.1.5
|
| [51] |
P. Mutsuddy, S. Tahmina Jhora, A. K. M. Shamsuzzaman, S. M. G. Kaisar, M. N. A. Khan, S. Dhiman, Dengue situation in Bangladesh: An epidemiological shift in terms of morbidity and mortality, Canadian J. Infect. Dis. Med. Microbiol., 2019 (2019), 3516284. https://doi.org/10.1155/2019/3516284 doi: 10.1155/2019/3516284
|
| [52] |
A. Chamnan, P. Pongsumpun, I. M. Tang, N. Wongvanich, Local and global stability analysis of dengue disease with vaccination and optimal control, Symmetry, 13 (2021), 1917. https://doi.org/10.3390/sym13101917 doi: 10.3390/sym13101917
|
| [53] |
H. S. Rodrigues, M. T. T. Monteiro, D. F. M. Torres, Sensitivity analysis in a dengue epidemiological model, Conference Papers in Mathematics, 2013 (2013), 721406. https://doi.org/10.1155/2013/721406 doi: 10.1155/2013/721406
|
| [54] |
M. R. Hasan, A. Hobiny, A. Alshehri, Analysis of vector-host SEIR-SEI dengue epidemiological model, Int. J. Anal. Appl., 20 (2022), 57. https://doi.org/10.28924/2291-8639-20-2022-57 doi: 10.28924/2291-8639-20-2022-57
|
| [55] |
A. Omame, M. E. Isah, M. Abbas, An optimal control model for COVID‐19, zika, dengue, and chikungunya co‐dynamics with reinfection, Optim. Control Appl. Method., 44 (2022), 170–204. https://doi.org/10.1002/oca.2936 doi: 10.1002/oca.2936
|
| [56] | A. Dwivedi, V. Baniya, R. Keval, Application of optimal controls on dengue dynamics-A mathematical study, In: Mathematical modelling and computational intelligence techniques, Springer, 2021. https://doi.org/10.1007/978-981-16-6018-4_1 |








Figures(9) / Tables(3)

Turki D. Alharbi, Md Rifat Hasan. Global stability and sensitivity analysis of vector-host dengue mathematical model[J]. AIMS Mathematics, 2024, 9(11): 32797-32818. doi: 10.3934/math.20241569







 DownLoad:
DownLoad: