Resolving set has several applications in the fields of science, engineering, and computer science. One application of the resolving set problem includes navigation robots, chemical structures, and supply chain management. Suppose the set $ W = \left\{{s}_{1}, {s}_{2}, \dots , {s}_{k}\right\}\subset V\left(G\right) $, the vertex representations of $ x\in V\left(G\right) $ is $ {r}_{m}\left(x\right|W) = \{d(x, {s}_{1}), d(x, {s}_{2}), \dots , d(x, {s}_{k})\} $, where $ d(x, {s}_{i}) $ is the length of the shortest path of the vertex $ x $ and the vertex in $ W $ together with their multiplicity. The set $ W $ is called a local $ m $-resolving set of graphs $ G $ if $ {r}_{m}\left(v|W\right)\ne {r}_{m}\left(u\right|W) $ for $ uv\in E\left(G\right) $. The local $ m $-resolving set having minimum cardinality is called the local multiset basis and its cardinality is called the local multiset dimension of $ G $, denoted by $ m{d}_{l}\left(G\right) $. In our paper, we determined the bounds of the local multiset dimension of the comb product of tree graphs.
Citation: Ridho Alfarisi, Liliek Susilowati, Dafik. Local multiset dimension of comb product of tree graphs[J]. AIMS Mathematics, 2023, 8(4): 8349-8364. doi: 10.3934/math.2023421
Resolving set has several applications in the fields of science, engineering, and computer science. One application of the resolving set problem includes navigation robots, chemical structures, and supply chain management. Suppose the set $ W = \left\{{s}_{1}, {s}_{2}, \dots , {s}_{k}\right\}\subset V\left(G\right) $, the vertex representations of $ x\in V\left(G\right) $ is $ {r}_{m}\left(x\right|W) = \{d(x, {s}_{1}), d(x, {s}_{2}), \dots , d(x, {s}_{k})\} $, where $ d(x, {s}_{i}) $ is the length of the shortest path of the vertex $ x $ and the vertex in $ W $ together with their multiplicity. The set $ W $ is called a local $ m $-resolving set of graphs $ G $ if $ {r}_{m}\left(v|W\right)\ne {r}_{m}\left(u\right|W) $ for $ uv\in E\left(G\right) $. The local $ m $-resolving set having minimum cardinality is called the local multiset basis and its cardinality is called the local multiset dimension of $ G $, denoted by $ m{d}_{l}\left(G\right) $. In our paper, we determined the bounds of the local multiset dimension of the comb product of tree graphs.

| [1] | S. Khuller, B. Raghavachari, A. Rosenfeld, Localization in graphs, 1994. Available from: http://hdl.handle.net/1903/655. |
| [2] |
G. Chartrand, L. Eroh, M. A. Johnson, O. R. Oellermann, Resolvability in graphs and the metric dimension of a graph, Discrete Appl. Math., 105 (2000), 99–113. https://doi.org/10.1016/S0166-218X(00)00198-0 doi: 10.1016/S0166-218X(00)00198-0

|
| [3] | F. Okamoto, B. Phinezy, P. Zhang, The Local metric dimension of a graph, Math. Bohem., 135 (2010), 239–255. |
| [4] | R. Simanjuntak, P. Siagian, T. Vetrik, The multiset dimension of graphs, 2017. Available from: https://doi.org/10.48550/arXiv.1711.00225. |
| [5] | R. Alfarisi, Dafik, A. I. Kristiana, I. H. Agustin, The local multiset dimension of graphs, IJET 8 (2019), 120–124. |
| [6] |
R. Alfarisi, Y. Lin, J. Ryan, Dafik, I. H. Agustin, A note on multiset dimension and local multiset dimension of graphs, Stat., Optim. & Inf. Comput., 8 (2020), 890–901. https://doi.org/10.19139/soic-2310-5070-727 doi: 10.19139/soic-2310-5070-727
|
| [7] |
R. Adawiyah, Dafik, I. H. Agustin, R. M. Prihandini, R. Alfarisi, E. R. Albirri, On the local multiset dimension of an m-shadow graph, J. Phys.: Conf. Ser., 1211 (2019), 012006. https://doi.org/10.1088/1742-6596/1211/1/012006 doi: 10.1088/1742-6596/1211/1/012006
|
| [8] |
R. Alfarisi, M. I. Utoyo, Dafik, Local multiset dimension of related cycle graphs, AIP Conf. Proc., 2391 (2022), 080008. https://doi.org/10.1063/5.0072516 doi: 10.1063/5.0072516
|
| [9] |
R. Adawiyah, R. M. Prihandini, E. R. Albirri, Dafik, I. H. Agustin, R. Alfarisi, The local multiset dimension of a unicyclic graph, IOP Conf. Ser.: Earth Environ. Sci., 243 (2019), 012075. https://doi.org/10.1088/1755-1315/243/1/012075 doi: 10.1088/1755-1315/243/1/012075
|
| [10] | H. Iswadi, E. T. Baskoro, A. N. M. Salman, R. Simanjuntak, The resolving graph of amalgamation of cycles, Utilitas Math., 83 (2010), 121–132. |
| [11] | R. Diestel, Graph theory, Heidelberg: Springer, 2016. |
| [12] | R. Alfarisi, L. Susilowati, Dafik, The Local multiset resolving of graphs, 2022, In press. |
| [13] | S. W. Saputro, N. Mardiana, I. A. Purwasih, The metric dimension of comb product graph, Mat. Vestn., 4 (2017), 248–258. |
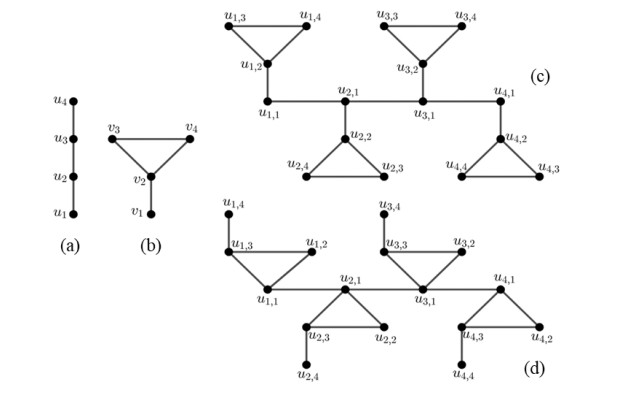
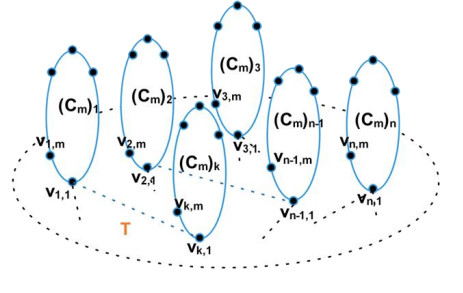
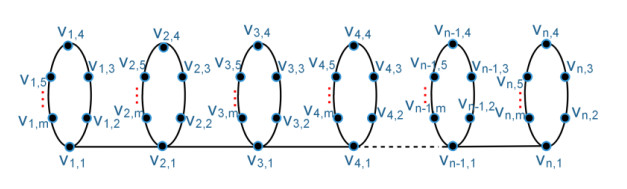
Figures(3) / Tables(1)

Ridho Alfarisi, Liliek Susilowati, Dafik. Local multiset dimension of comb product of tree graphs[J]. AIMS Mathematics, 2023, 8(4): 8349-8364. doi: 10.3934/math.2023421







 DownLoad:
DownLoad: