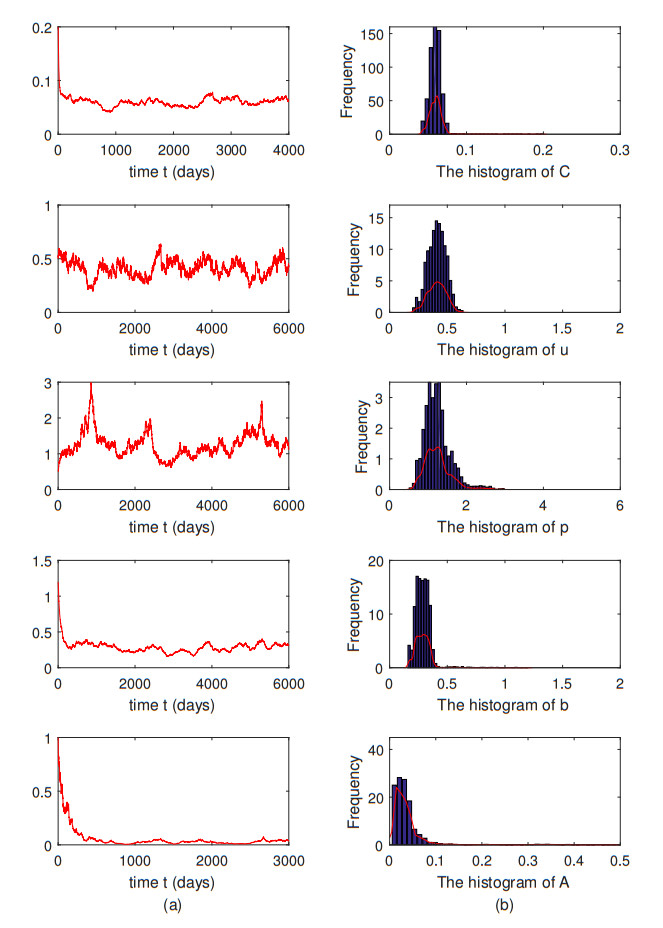
In this paper, a stochastic Alzheimer's disease model with the effect of calcium on amyloid beta is proposed. The Lyapunov function is constructed, followed by the feasibility and positivity and the existence of a stationary distribution for the positive solutions of the proposed model. The sufficient conditions for the extinction of the stochastic Alzheimer's disease model are derived through the Lyapunov function. This indicates that beta-amyloid plaque and the complex of beta-amyloid oligomers with prion protein may go extinct and there is a possibility of a cure for the disease. Furthermore, our numerical simulations show that as the intensity of the random disturbance increases, the time it takes for the disease to go extinct decreases.
Citation: Ruoyun Lang, Yuanshun Tan, Yu Mu. Stationary distribution and extinction of a stochastic Alzheimer's disease model[J]. AIMS Mathematics, 2023, 8(10): 23313-23335. doi: 10.3934/math.20231185
In this paper, a stochastic Alzheimer's disease model with the effect of calcium on amyloid beta is proposed. The Lyapunov function is constructed, followed by the feasibility and positivity and the existence of a stationary distribution for the positive solutions of the proposed model. The sufficient conditions for the extinction of the stochastic Alzheimer's disease model are derived through the Lyapunov function. This indicates that beta-amyloid plaque and the complex of beta-amyloid oligomers with prion protein may go extinct and there is a possibility of a cure for the disease. Furthermore, our numerical simulations show that as the intensity of the random disturbance increases, the time it takes for the disease to go extinct decreases.

| [1] |
S. Jean-Marie, Z. Jadwiga, J. Pierre, A possible role for CSF turnover and choroid plexus in the pathogenesis of late onset Alzheimer's disease, J. Alzheimer's Dis., 30 (2012), 17–26. https://doi.org/10.3233/JAD-2012-111964 doi: 10.3233/JAD-2012-111964

|
| [2] |
R. Bai, W. Dong, Trends in mortality rates for Alzheimer's disease and other dementias over 30 years in China, Am. J. Alzheimer's Dis., 36 (2021), 1–8. https://doi.org/10.1177/15333175211044884 doi: 10.1177/15333175211044884
|
| [3] |
Alzheimer's Association Report, 2021 Alzheimer's disease facts and figures, Alzheimer's Dement., 17 (2021), 327–406. https://doi.org/10.1002/alz.12328 doi: 10.1002/alz.12328
|
| [4] |
L. Söderberg, M. Johannesson, P. Nygren, H. Laudon, F. Eriksson, G. Osswald, et al., Lecanemab, Aducanumab, and Gantenerumab–Binding profiles to different forms of amyloid-beta might explain efficacy and side effects in clinical trials for Alzheimer's disease, Neurotherapeutics, 20 (2023), 195–206. https://doi.org/10.1007/s13311-022-01308-6 doi: 10.1007/s13311-022-01308-6
|
| [5] |
J. Cummings, P. Aisen, C. Lemere, A. Atri, M. Sabbagh, S. Salloway, Aducanumab produced a clinically meaningful benefit in association with amyloid lowering, Alz. Res. Therapy, 13 (2021), 98. https://doi.org/10.1186/s13195-021-00838-z doi: 10.1186/s13195-021-00838-z
|
| [6] |
K. Wojtunik-Kulesza, M. Rudkowska, A. Orzeł-Sajdłowska, Aducanumab-Hope or disappointment for Alzheimer's disease, Int. J. Mol. Sci., 24 (2023), 4367. https://doi.org/10.3390/ijms24054367 doi: 10.3390/ijms24054367
|
| [7] |
Y. Huang, L. Mucke, Alzheimer mechanisms and therapeutic strategies, Cell, 148 (2012), 1204–1222. https://doi.org/10.1016/j.cell.2012.02.040 doi: 10.1016/j.cell.2012.02.040
|
| [8] |
M. P. Lambert, A. K. Barlow, B. A. Chromy, W. L. Klein, Diffusible, nonfibrillar ligands derived from $ A\beta_{1-42}$ are potent central nervous system neurotoxins, Proc. Natl. Acad. Sci. USA, 95 (1998), 6448–6453. https://doi.org/10.1073/pnas.95.11.6448 doi: 10.1073/pnas.95.11.6448
|
| [9] |
M. Hoore, S. Khailaie, G. Montaseri, T. Mitra, M. Meyer-Hermann, Mathematical model shows how sleep may affect amyloid-$\beta$ fibrillization, Biophys. J., 119 (2018), 862–872. https://doi.org/10.1016/j.bpj.2020.07.011 doi: 10.1016/j.bpj.2020.07.011
|
| [10] |
H. Li, H. Zhao, Mathematical model of Alzheimer's disease with prion proteins interactions and treatment, Appl. Math. Comput., 433 (2022), 127377. https://doi.org/10.1016/j.amc.2022.127377 doi: 10.1016/j.amc.2022.127377
|
| [11] |
F. Dohler, D. Sepulveda-Falla, S. Krasemann, H. Altmeppen, H. Schlüter, D. Hildebrand, et al., High molecular mass assemblies of amyloid-$\beta$ oligomers bind prion protein in patients with Alzheimer's disease, Brain, 137 (2014), 873–886. https://doi.org/10.1093/brain/awt375 doi: 10.1093/brain/awt375
|
| [12] |
B. Mroczko, M. Groblewska, A. Litman-Zawadzka, J. Kornhuber, P. Lewczuk, Amyloid $\beta$ oligomers (A$\beta$Os) in Alzheimer's disease, J. Neural Transm., 125 (2018), 177–191. https://doi.org/10.1007/s00702-017-1820-x doi: 10.1007/s00702-017-1820-x
|
| [13] |
M. Helal, E. Hingant, L. Pujo-Menjouet, G. F. Webb, Alzheimer's disease: analysis of a mathematical model incorporating the role of prions, J. Math. Biol., 69 (2014), 1207–1235. https://doi.org/10.1007/s00285-013-0732-0 doi: 10.1007/s00285-013-0732-0
|
| [14] |
J. Hu, Q. Zhang, A. Meyer-Baese, M. Ye, Finite-time stability and optimal control of a stochastic reaction-diffusion model for Alzheimer's disease with impulse and time-varying delay, Appl. Math. Model., 102 (2022), 511–539. https://doi.org/10.1016/j.apm.2021.10.004 doi: 10.1016/j.apm.2021.10.004
|
| [15] |
B. Fang, D. Wang, M. Huang, G. Yu, H. Li, Hypothesis on the relationship between the change in intracellular pH and incidence of sporadic Alzheimer's disease or vascular dementia, Int. J. Neurosci. 120 (2010), 591–595. https://doi.org/10.3109/00207454.2010.505353 doi: 10.3109/00207454.2010.505353
|
| [16] |
A. De Mario, C. Peggion, M. L. Massimino, F. Viviani, A. Castellani, M. Giacomello, et al., The prion protein regulates glutamate-mediated $ Ca^{2+} $ entry and mitochondrial $ Ca^{2+} $ accumulation in neurons, J. Cell Sci., 130 (2017), 2736–2746. https://doi.org/10.1242/jcs.196972 doi: 10.1242/jcs.196972
|
| [17] |
H. J. Kwon, M. Y. Cha, D. Kim, D. K. Kim, M. Soh, K. Shin, et al., Mitochondria-targeting ceria nanoparticles as antioxidants for Alzheimer's disease, ACS Nano, 10 (2016), 2860–2870. https://doi.org/10.1021/acsnano.5b08045 doi: 10.1021/acsnano.5b08045
|
| [18] |
A. Pannaccione, I. Piccialli, A. Secondo, R. Ciccone, P. Molinaro, F. Boscia, et al., The $ Na^{+} $/$ Ca^{2+} $exchanger in Alzheimer's disease, Cell Calcium, 87 (2020), 102190. https://doi.org/10.1016/j.ceca.2020.102190 doi: 10.1016/j.ceca.2020.102190
|
| [19] |
J. De Caluwé, G. Dupont, The progression towards Alzheimer's disease described as a bistable switch arising from the positive loop between amyloids and $ Ca^{2+} $, J. Theor. Biol., 331 (2013), 12–18. https://doi.org/10.1016/j.jtbi.2013.04.015 doi: 10.1016/j.jtbi.2013.04.015
|
| [20] | B. Øksendal, Stochastic differential equations: an introduction with applications, Heidelberg: Springer Berlin, 2003. https://doi.org/10.1007/978-3-642-14394-6 |
| [21] |
M. Xin, B. Wang, Stationary distribution and extinction of a stochastic tuberculosis model, Phys. A, 545 (2020), 123741. https://doi.org/10.1016/j.physa.2019.123741 doi: 10.1016/j.physa.2019.123741
|
| [22] |
M. Liu, M. Deng, Permanence and extinction of a stochastic hybrid model for tumor growth, Appl. Math. Lett., 94 (2019), 66–72. https://doi.org/10.1016/j.aml.2019.02.016 doi: 10.1016/j.aml.2019.02.016
|
| [23] |
Y. Tan, Y. Cai, X. Wang, Z. Peng, K. Wang, R. Yao, et al., Stochastic dynamics of an SIS epidemiological model with media coverage, Math. Comput. Simul., 204 (2023), 1–27. https://doi.org/10.1016/j.matcom.2022.08.001 doi: 10.1016/j.matcom.2022.08.001
|
| [24] |
J. Hu, Q. Zhang, A. Meyer-Baese, M. Ye, Stationary distribution of a stochastic Alzheimer's disease model, Math. Methods Appl. Sci., 43 (2020), 9706–9718. https://doi.org/10.1002/mma.6642 doi: 10.1002/mma.6642
|
| [25] |
T. Matsubara, K. Satoh, T. Homma, T. Nakagaki, N. Yamaguchi, R. Atarashi, et al., Prion protein interacts with the metabotropic glutamate receptor 1 and regulates the organization of $ Ca^{2+} $ signaling, Biochem. Biophys. Res. Commun., 525 (2020), 447–454. https://doi.org/10.1016/j.bbrc.2020.02.102 doi: 10.1016/j.bbrc.2020.02.102
|
| [26] |
J. Latulippe, D. Lotito, D. Murby, A mathematical model for the effects of amyloid beta on intracellular calcium, PLoS One, 13 (2018), 1–27. https://doi.org/10.1371/journal.pone.0202503 doi: 10.1371/journal.pone.0202503
|
| [27] |
A. Din, T. Khan, Y. Li, H. Tahir, A. Khan, W. A. Khan, Mathematical analysis of dengue stochastic epidemic model, Results Phys., 20 (2021), 103719. https://doi.org/10.1016/j.rinp.2020.103719 doi: 10.1016/j.rinp.2020.103719
|
| [28] |
W. Wang, A. Zazoua, Analysis of mathematical model of prostate cancer with androgen deprivation therapy, Commun. Nonlinear Sci. Numer. Simul., 66 (2019), 41–60. https://doi.org/10.1016/j.cnsns.2018.06.004 doi: 10.1016/j.cnsns.2018.06.004
|
| [29] |
Y. Wang, D. Jiang, Stationary distribution and extinction of a stochastic viral infection model, Discrete Dyn. Nat. Soc., 2017 (2017), 6027509. https://doi.org/10.1155/2017/6027509 doi: 10.1155/2017/6027509
|
| [30] | R. Khasminskii, Stochastic stability of differential equations, Heidelberg: Springer Berlin, 2012. https://doi.org/10.1007/978-3-642-23280-0 |
| [31] |
X. Mao, C. Yuan, J. Zou, Stochastic differential delay equations of population dynamics, J. Math. Anal. Appl., 304 (2005), 296–320. https://doi.org/10.1016/j.jmaa.2004.09.027 doi: 10.1016/j.jmaa.2004.09.027
|
| [32] |
G. Lan, Z. Lin, C. Wei, S. Zhang, A stochastic SIRS epidemic model with non-monotone incidence rate under regime-switching, J. Franklin Inst., 16 (2019), 9844–9866. https://doi.org/10.1016/j.jfranklin.2019.09.009 doi: 10.1016/j.jfranklin.2019.09.009
|
| [33] |
J. Olazarán, B. Reisberg, L. Clare, I. Cruz, J. Peña-Casanova, T. del Ser, et al., Nonpharmacological therapies in Alzheimer's disease: a systematic review of efficacy, Dement. Geriatr. Cogn. Disord., 30 (2010), 161–178. https://doi.org/10.1159/000316119 doi: 10.1159/000316119
|
| [34] |
D. J. Higham, An algorithmic introduction to numerical simulation of stochastic differential equations, SIAM Rev., 43 (2001), 525–546. https://doi.org/10.1137/S0036144500378302 doi: 10.1137/S0036144500378302
|


Figures(3)

Ruoyun Lang, Yuanshun Tan, Yu Mu. Stationary distribution and extinction of a stochastic Alzheimer's disease model[J]. AIMS Mathematics, 2023, 8(10): 23313-23335. doi: 10.3934/math.20231185







 DownLoad:
DownLoad: