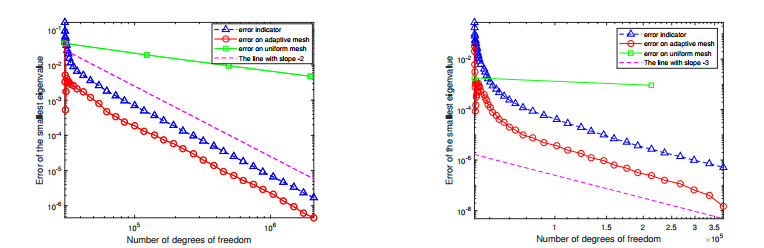
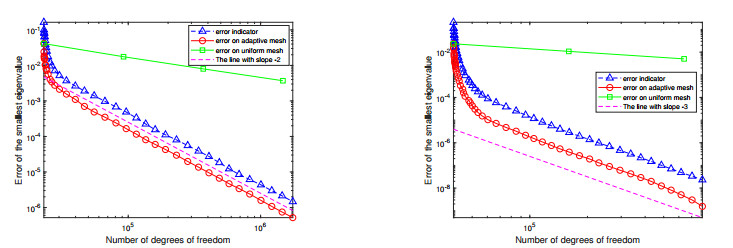
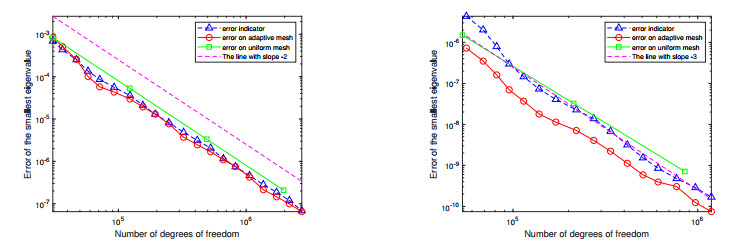
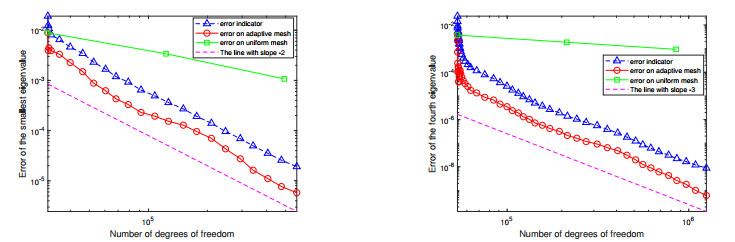
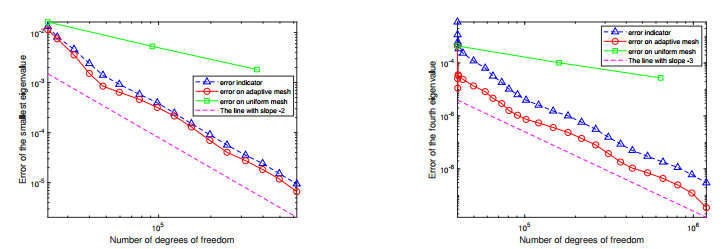
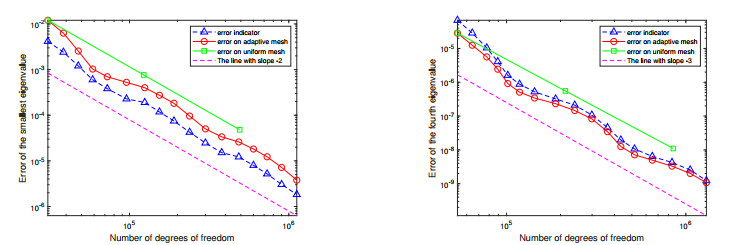
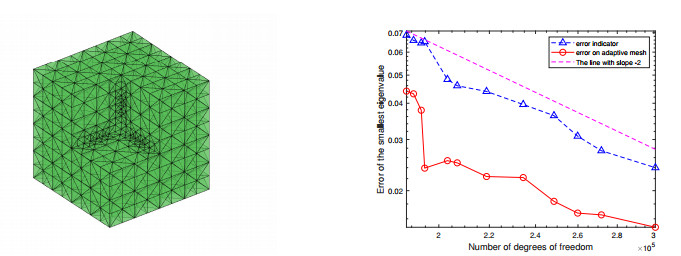
For a class of Stokes eigenvalue problems including the classical Stokes eigenvalue problem and the magnetohydrodynamic Stokes eigenvalue problem a residual type a posteriori error estimate of the mixed discontinuous Galerkin finite element method using $ \mathbb{P}_{k}-\mathbb{P}_{k-1} $ element $ (k\geq 1) $ is studied in this paper. The a posteriori error estimators for approximate eigenpairs are given. The reliability and efficiency of the posteriori error estimator for the eigenfunction are proved and the reliability of the estimator for the eigenvalue is also analyzed. The numerical results are provided to confirm the theoretical predictions and indicate that the method considered in this paper can reach the optimal convergence order $ O(dof^{\frac{-2k}{d}}) $.
Citation: Lingling Sun, Hai Bi, Yidu Yang. A posteriori error estimates of mixed discontinuous Galerkin method for a class of Stokes eigenvalue problems[J]. AIMS Mathematics, 2023, 8(9): 21270-21297. doi: 10.3934/math.20231084
For a class of Stokes eigenvalue problems including the classical Stokes eigenvalue problem and the magnetohydrodynamic Stokes eigenvalue problem a residual type a posteriori error estimate of the mixed discontinuous Galerkin finite element method using $ \mathbb{P}_{k}-\mathbb{P}_{k-1} $ element $ (k\geq 1) $ is studied in this paper. The a posteriori error estimators for approximate eigenpairs are given. The reliability and efficiency of the posteriori error estimator for the eigenfunction are proved and the reliability of the estimator for the eigenvalue is also analyzed. The numerical results are provided to confirm the theoretical predictions and indicate that the method considered in this paper can reach the optimal convergence order $ O(dof^{\frac{-2k}{d}}) $.

| [1] |
J. Gedicke, A. Khan, Arnold-Winther mixed finite elements for Stokes eigenvalue problems, SIAM J. Sci. Comput., 40 (2018), A3449–A3469. https://doi.org/10.1137/17M1162032 doi: 10.1137/17M1162032

|
| [2] |
J. Gedicke, A. Khan, Divergence-conforming discontinuous Galerkin finite elements for Stokes eigenvalue problems, Numer. Math., 144 (2020), 585–614. https://doi.org/10.1007/s00211-019-01095-x doi: 10.1007/s00211-019-01095-x
|
| [3] |
F. Lepea, D. Mora, Symmetric and nonsymmetric discontinuous Galerkin methods for a pseudostress formulation of the Stokes spectral problem, SIAM J. Sci. Comput., 42 (2020), A698–A722. https://doi.org/10.1137/19M1259535 doi: 10.1137/19M1259535
|
| [4] |
F. Lepe, G. Rivera, A virtual element approximation for the pseudostress formulation of the Stokes eigenvalue problem, Comput. Methods Appl. Mech. Eng., 379 (2021), 113753. https://doi.org/10.1016/j.cma.2021.113753 doi: 10.1016/j.cma.2021.113753
|
| [5] |
B. Mercier, J. Osborn, J. Rappaz, P. A. Raviart, Eigenvalue approximation by mixed and hybrid methods, Math. Comp., 36 (1981), 427–453. https://doi.org/10.2307/2007651 doi: 10.2307/2007651
|
| [6] |
L. L. Sun, Y. D. Yang, The a posteriori error estimates and adaptive computation of nonconforming mixed finite elements for the Stokes eigenvalue problem, Appl. Math. Comput., 421 (2022), 126951. https://doi.org/10.1016/j.amc.2022.126951 doi: 10.1016/j.amc.2022.126951
|
| [7] |
L. L. Sun, H. Bi, Y. D. Yang, The discontinuous Galerkin and the nonconforming ECR element approximations for an MHD Stokes eigenvalue problem, Math. Methods Appl. Sci., 46 (2022), 6154–6176. https://doi.org/10.1002/mma.8897 doi: 10.1002/mma.8897
|
| [8] |
Ö. Türk, An MHD Stokes eigenvalue problem and its approximation by a spectral collocation method, Comput. Math. Appl., 80 (2020), 2045–2056. https://doi.org/10.1016/j.camwa.2020.09.002 doi: 10.1016/j.camwa.2020.09.002
|
| [9] |
M. G. Armentano, V. Moreno, A posteriori error estimates of stabilized low-order mixed finite elements for the Stokes eigenvalue problem, J. Comput. Appl. Math., 269 (2014), 132–149. https://doi.org/10.1016/j.cam.2014.03.027 doi: 10.1016/j.cam.2014.03.027
|
| [10] |
H. P. Liu, W. Gong, S. H. Wang, N. N. Yan, Superconvergence and a posteriori error estimates for the Stokes eigenvalue problems, BIT Numer. Math., 53 (2013), 665–687. https://doi.org/10.1007/s10543-013-0422-8 doi: 10.1007/s10543-013-0422-8
|
| [11] |
C. Lovadina, M. Lyly, R. Stenberg, A posteriori estimates for the Stokes eigenvalue problem, Numer. Methods Partial Differ. Equ., 25 (2009), 244–257. https://doi.org/10.1002/num.20342 doi: 10.1002/num.20342
|
| [12] |
S. H. Jia, F. S. Lu, H. H. Xie, A Posterior error analysis for the nonconforming discretization of Stokes eigenvalue problem, Acta Math. Sin. Engl. Ser., 30 (2014), 949–967. https://doi.org/10.1007/s10114-014-3121-8 doi: 10.1007/s10114-014-3121-8
|
| [13] |
F. Lepe, G. Rivera, J. Vellojin, Error estimates for a vorticity-based velocity-stress formulation of the Stokes eigenvalue problem, J. Comput. Appl. Math., 420 (2023), 114798. https://doi.org/10.1016/j.cam.2022.114798 doi: 10.1016/j.cam.2022.114798
|
| [14] |
S. Badia, R. Codina, T. Gudi, J. Guzmán, Error analysis of discontinuous Galerkin methods for the Stokes problem under minimal regularity, IMA. J. Numer. Anal., 34 (2014), 800–819. https://doi.org/10.1093/imanum/drt022 doi: 10.1093/imanum/drt022
|
| [15] | B. Cockburn, G. E. Karniadakis, C. W. Shu, Discontinuous Galerkin methods: theory, computation and applications, Berlin, Heidelberg: Springer, 2000. https://doi.org/10.1007/978-3-642-59721-3 |
| [16] | A. Ern, J. Guermond, Finite elements Ⅱ: Galerkin approximation, elliptic and mixed PDEs, Cham: Springer, 2021. https://doi.org/10.1007/978-3-030-56923-5 |
| [17] |
P. Houston, D. Schötzau, T. P. Wihler, Energy norm a posteriori error estimation for mixed discontinuous Galerkin approximations of the Stokes problem, J. Sci. Comput., 22 (2005), 347–370. https://doi.org/10.1007/s10915-004-4143-7 doi: 10.1007/s10915-004-4143-7
|
| [18] | P. Houston, D. Schötzau, T. P. Wihler, $hp$-adaptive discontinuous Galerkin finite element methods for the Stokes problem, European Congress on Computational Methods in Applied Sciences and Engineering (ECCOMAS), 2004, 1–20. |
| [19] |
D. A. Di Pietro, A. Ern, Discrete functional analysis tools for discontinuous Galerkin methods with application to the incompressible Navier-Stokes equations, Math. Comp., 79 (2010), 1303–1330. https://doi.org/10.1090/s0025-5718-10-02333-1 doi: 10.1090/s0025-5718-10-02333-1
|
| [20] | M. Ainsworth, J. T. Oden, A posteriori error estimation in finite element analysis, New York: Wiley, 2011. |
| [21] | R. Verfürth, A posteriori error estimation techniques, New York: Oxford University Press, 1996. |
| [22] |
S. C. Brenner, Poincaré-Friedrichs inequalities for piecewise $H^{1}$ functions, SIAM J. Numer. Anal., 41 (2003), 306–324. https://doi.org/10.1137/S0036142902401311 doi: 10.1137/S0036142902401311
|
| [23] |
O. A. Karakashian, F. Pascal, A posteriori error estimates for a discontinuous Galerkin approximation of second-order elliptic problems, SIAM J. Numer. Anal., 41 (2003), 2374–2399. https://doi.org/10.1137/s0036142902405217 doi: 10.1137/s0036142902405217
|
| [24] |
I. Perugia, D. Schötzau, The $hp$-local discontinuous Galerkin method for low-frequency time-harmonic Maxwell equations, Math. Comp., 72 (2003), 1179–1214. https://doi.org/10.1090/s0025-5718-02-01471-0 doi: 10.1090/s0025-5718-02-01471-0
|
| [25] |
D. Schötzau, C. Schwab, A. Toselli, Mixed $hp$-DGFEM for incompressible flows, SIAM J. Numer. Anal., 40 (2002), 2171–2194. https://doi.org/10.1137/s0036142901399124 doi: 10.1137/s0036142901399124
|
| [26] |
P. Houston, I. Perugia, D. Schötzau, An a posteriori error indicator for discontinuous Galerkin discretizations of H(curl)-elliptic partial differential equations, IMA J. Numer. Anal., 27 (2007), 122–150. https://doi.org/10.1093/imanum/drl012 doi: 10.1093/imanum/drl012
|
| [27] | B. Rivière, Discontinuous Galerkin methods for solving elliptic and parabolic equations: theory and implementation, SIAM, 2008. |
| [28] | V. Girault, P. A. Raviart, Finite element approximation of the Navier-Stokes equations, Berlin, Heidelberg: Springer, 1979. |
| [29] |
B. Cockburn, G. Kanschat, D. Schötzau, C. Schwab, Local discontinuous Galerkin methods for the Stokes system, SIAM J. Numer. Anal., 40 (2002), 319–343. https://doi.org/10.1137/S0036142900380121 doi: 10.1137/S0036142900380121
|
| [30] |
P. Hansbo, M. G. Larson, Discontinuous Galerkin methods for incompressible and nearly incompressible elasticity by Nitsche's method, Comput. Methods Appl. Mech. Eng., 191 (2002), 1895–1908. https://doi.org/10.1016/S0045-7825(01)00358-9 doi: 10.1016/S0045-7825(01)00358-9
|
| [31] |
Y. D. Yang, Z. M. Zhang, F. B. Lin, Eigenvalue approximation from below using non-conforming finite elements, Sci. China Ser. A Math., 53 (2010), 137–150. https://doi.org/10.1007/s11425-009-0198-0 doi: 10.1007/s11425-009-0198-0
|
| [32] | I. Babuska, J. Osborn, Eigenvalue problems, In: Handbook of numerical analysis, Elsevier, 1991,641–787. https://doi.org/10.1016/S1570-8659(05)80042-0 |
| [33] |
D. Boffi, Finite element approximation of eigenvalue problems, Acta Numer., 19 (2010), 1–120. https://doi.org/10.1017/S0962492910000012 doi: 10.1017/S0962492910000012
|
| [34] |
L. R. Scott, S. Y. Zhang, Finite element interpolation of nonsmooth functions satisfying boundary conditions, Math. Comp., 54 (1990), 483–493. https://doi.org/10.1090/S0025-5718-1990-1011446-7 doi: 10.1090/S0025-5718-1990-1011446-7
|
| [35] |
G. Kanschat, D. Schötzau, Energy norm a posteriori error estimation for divergence-free discontinuous Galerkin approximations of the Navier-Stokes equations, Int. J. Numer. Methods Fluids, 57 (2008), 1093–1113. https://doi.org/10.1002/fld.1795 doi: 10.1002/fld.1795
|
| [36] |
Y. P. Zeng, F. Wang, A posteriori error estimates for a discontinuous Galerkin approximation of Steklov eigenvalue problems, Appl. Math., 62 (2017), 243–267. https://doi.org/10.21136/AM.2017.0115-16 doi: 10.21136/AM.2017.0115-16
|
| [37] |
H. J. Wu, Z. M. Zhang, Can we have superconvergent gradient recovery under adaptive meshes? SIAM J. Numer. Anal., 45 (2007), 1701–1722. https://doi.org/10.1137/060661430 doi: 10.1137/060661430
|
| [38] |
Y. D. Yang, Y. Zhang, H. Bi, A type of adaptive $C^{0}$ non-conforming finite element method for the Helmholtz transmission eigenvalue problem, Comput. Methods Appl. Mech. Eng., 360 (2020), 112697. https://doi.org/10.1016/j.cma.2019.112697 doi: 10.1016/j.cma.2019.112697
|
| [39] |
Y. J. Li, H. Bi, Y. D. Yang, The a priori and a posteriori error estimates of DG method for the Steklov eigenvalue problem in inverse scattering, J. Sci. Comput., 91 (2022), 20. https://doi.org/10.1007/s10915-022-01787-x doi: 10.1007/s10915-022-01787-x
|
| [40] |
X. Y. Dai, J. C. Xu, A. H. Zhou, Convergence and optimal complexity of adaptive finite element eigenvalue computations, Numer. Math., 110 (2008), 313–355. https://doi.org/10.1007/s00211-008-0169-3 doi: 10.1007/s00211-008-0169-3
|
| [41] |
W. Dörfler, A convergent adaptive algorithm for Poisson's equation, SIAM J. Numer. Anal., 33 (1996), 1106–1124. https://doi.org/10.1137/0733054 doi: 10.1137/0733054
|
| [42] |
P. Morin, R. H. Nochetto, K. G. Siebert, Convergence of adaptive finite element methods, SIAM Rev., 44 (2002), 631–658. https://doi.org/10.1137/s0036144502409093 doi: 10.1137/s0036144502409093
|
| [43] | L. Chen, iFEM: An integrated finite element method package in Matlab, Technical Report, University of California at Irvine, 2009. |
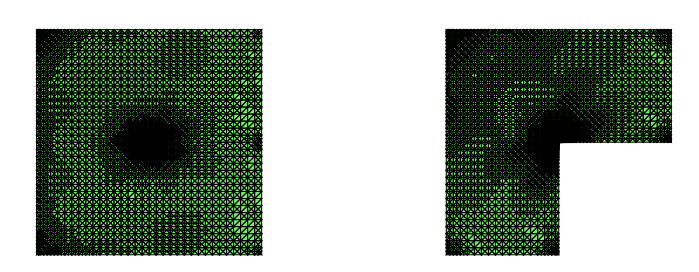

Figures(12) / Tables(8)

Lingling Sun, Hai Bi, Yidu Yang. A posteriori error estimates of mixed discontinuous Galerkin method for a class of Stokes eigenvalue problems[J]. AIMS Mathematics, 2023, 8(9): 21270-21297. doi: 10.3934/math.20231084







 DownLoad:
DownLoad: