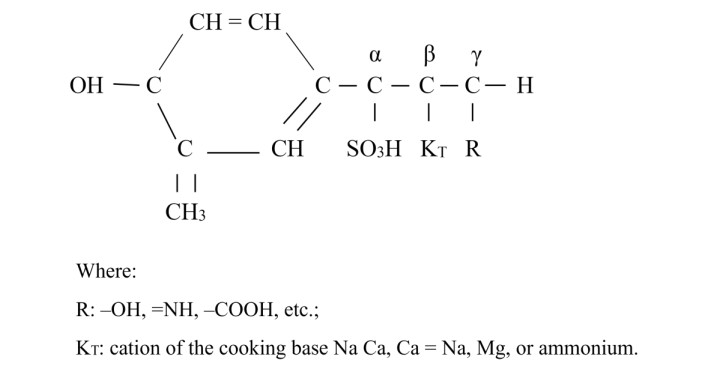
This study proves that lignin-based biopolymer materials can be employed as starting materials for the synthesis of novel casting binders that fulfill the current level of characteristics. The optimal concentration of the binder in the mixture was experimentally determined to be 5.8%–6.2%. It has been demonstrated in practice that the employment of ammonium salts as a technical lignosulfonate (TLS) modifier can result in the provision of cold (room temperature) curing of a mixture based on them. It was proposed to use as a technological additive that boosts the strength characteristics of a mixture of substances carboxymethyl cellulose (CMC). In a variety of adhesive materials, it is utilized as an active polymer base. The concentration limits for using CMC in the mixture are set at 0.15%–0.25%. To improve the moldability of the combination, it was suggested that kaolin clay be used as a plasticizing addition. The concentration limits for using a plasticizing additive are set at 3.5%–4.0%. The produced mixture was compared to the analog of the alpha-set method in a comparative analysis. It was discovered that the proposed composition is less expensive, more environmentally friendly, and enables the production of high-quality castings. In terms of physical, mechanical, and technological properties, the created composition of the cold curing mixture is not inferior to analogs from the alpha-set method. For the first time, a biopolymer-based binder system containing technical lignosulfonate with the addition of ammonium sulfate and carboxymethyl cellulose was used in the production of cast iron castings on the case of a cylinder casting weighing 18.3 kg from gray cast iron grade SCh20. Thus, it has been proved possible for the first time to replace phenol-based resin binders with products based on natural polymer combinations. For the first time, a cold-hardening mixture based on technological lignosulfonates has been developed without using hardeners made of very hazardous and cancer-causing hexavalent chromium compounds. But is achieved through a combination of specialized additives, including kaolin clay to ensure the mixture can be manufactured, ammonium sulfate to ensure the mixture cures, and carboxymethyl cellulose to enhance the strength properties of the binder composition. The study's importance stems from the substitution of biopolymer natural materials for costly and environmentally harmful binders based on phenolic resins. This development's execution serves as an illustration of how green technology can be used in the foundry sector. Reducing the amount of resin used in foundry manufacturing and substituting it with biopolymer binders based on technological lignosulfonates results in lower product costs as well as the preservation of the environment. Using lignin products judiciously can reduce environmental harm by using technical lignosulfonates, or compounds based on technical lignin. The combination is concentrated on businesses with single and small-scale manufacturing because it is presumable that this is merely the beginning of the investigation. This study confirms the viability of creating a cold-hardening combination based on technical lignosulfonates in practical applications and supports this with the castings produced, using the creation of a gray cast iron cylinder casting as an example.
Citation: Falah Mustafa Al-Saraireh. Cold-curing mixtures based on biopolymer lignin complex for casting production in single and small-series conditions[J]. AIMS Materials Science, 2023, 10(5): 876-890. doi: 10.3934/matersci.2023047
This study proves that lignin-based biopolymer materials can be employed as starting materials for the synthesis of novel casting binders that fulfill the current level of characteristics. The optimal concentration of the binder in the mixture was experimentally determined to be 5.8%–6.2%. It has been demonstrated in practice that the employment of ammonium salts as a technical lignosulfonate (TLS) modifier can result in the provision of cold (room temperature) curing of a mixture based on them. It was proposed to use as a technological additive that boosts the strength characteristics of a mixture of substances carboxymethyl cellulose (CMC). In a variety of adhesive materials, it is utilized as an active polymer base. The concentration limits for using CMC in the mixture are set at 0.15%–0.25%. To improve the moldability of the combination, it was suggested that kaolin clay be used as a plasticizing addition. The concentration limits for using a plasticizing additive are set at 3.5%–4.0%. The produced mixture was compared to the analog of the alpha-set method in a comparative analysis. It was discovered that the proposed composition is less expensive, more environmentally friendly, and enables the production of high-quality castings. In terms of physical, mechanical, and technological properties, the created composition of the cold curing mixture is not inferior to analogs from the alpha-set method. For the first time, a biopolymer-based binder system containing technical lignosulfonate with the addition of ammonium sulfate and carboxymethyl cellulose was used in the production of cast iron castings on the case of a cylinder casting weighing 18.3 kg from gray cast iron grade SCh20. Thus, it has been proved possible for the first time to replace phenol-based resin binders with products based on natural polymer combinations. For the first time, a cold-hardening mixture based on technological lignosulfonates has been developed without using hardeners made of very hazardous and cancer-causing hexavalent chromium compounds. But is achieved through a combination of specialized additives, including kaolin clay to ensure the mixture can be manufactured, ammonium sulfate to ensure the mixture cures, and carboxymethyl cellulose to enhance the strength properties of the binder composition. The study's importance stems from the substitution of biopolymer natural materials for costly and environmentally harmful binders based on phenolic resins. This development's execution serves as an illustration of how green technology can be used in the foundry sector. Reducing the amount of resin used in foundry manufacturing and substituting it with biopolymer binders based on technological lignosulfonates results in lower product costs as well as the preservation of the environment. Using lignin products judiciously can reduce environmental harm by using technical lignosulfonates, or compounds based on technical lignin. The combination is concentrated on businesses with single and small-scale manufacturing because it is presumable that this is merely the beginning of the investigation. This study confirms the viability of creating a cold-hardening combination based on technical lignosulfonates in practical applications and supports this with the castings produced, using the creation of a gray cast iron cylinder casting as an example.

| [1] |
B. Pang, L. Lee, Opinion mining and sentiment analysis, Trends Inf. Retr., 2 (2008), 1–135. DOI: 10.1561/1500000011 doi: 10.1561/1500000011

|
| [2] |
G. Vinodhini, R. Chandrasekaran, Sentiment analysis and opinion mining: a survey, Int. J., 2 (2012), 282–292. DOI: 10.1016/j.nlp.2022.100003 doi: 10.1016/j.nlp.2022.100003
|
| [3] | M. Pontiki, D. Galanis, J. Pavlopoulos, H. Papageorgiou, S. Manandhar, SemEval-2014 Task 4: Aspect based sentiment analysis, in Association for Computational Linguistics, (2014), 27–35. DOI: 10.3115/v1/S14-2004 |
| [4] | M. Pontiki, D. Galanis, H. Papageorgiou, S. Manandhar, I. Androutsopoulos, Semeval-2015 task 12: Aspect based sentiment analysis, in Association for Computational Linguistics, (2015), 486–495. DOI: 10.18653/v1/S15-2082 |
| [5] | M. Pontiki, D. Galanis, H. Papageorgiou, I. Androutsopoulos, S. Manandhar, M. AL-Smadi, et al. Semeval-2016 task 5: Aspect based sentiment analysis, in Association for Computational Linguistics, (2016), 19–30. DOI: 10.18653/v1/S16-1002 |
| [6] | W. Zhang, X. Li, Y. Deng, L. Bing, W. Lam, A survey on aspect-based sentiment analysis: Tasks, methods, and challenges, IEEE Trans. Knowl. Data Eng., 2022. DOI: 10.1109/TKDE.2022.3230975 |
| [7] | D. Tang, B. Qin, X. Feng, T. Liu, Effective LSTMs for target-dependent sentiment classification, preprint, arXiv: 151201100. |
| [8] | M. Yang, W. Tu, J. Wang, F. Xu, X. Chen, Attention based LSTM for target dependent sentiment classification, in Proceedings of the AAAI conference on artificial intelligence, 2017. DOI: 10.1609/aaai.v31i1.11061 |
| [9] |
Q. Liu, Y. Huang, Q. Yang, H. Peng, J. Wang, An attention-aware long short-term memory-like spiking neural model for sentiment analysis, Int. J. Neural Syst., (2023), 2350037–2350037. DOI: 10.1142/s0129065723500375 doi: 10.1142/s0129065723500375
|
| [10] |
Y. Huang, Q. Liu, H. Peng, J. Wang, Q. Yang, D. Orellana-Martín, Sentiment classification using bidirectional LSTM-SNP model and attention mechanism, Expert Syst. Appl., 221 (2023), 119730. DOI: 10.1016/j.eswa.2023.119730 doi: 10.1016/j.eswa.2023.119730
|
| [11] |
Y. Huang, H. Peng, Q. Liu, Q. Yang, J. Wang, D. Orellana-Martín, et al., Attention-enabled gated spiking neural P model for aspect-level sentiment classification, Neural Network, 157 (2023), 437–443. DOI: 10.1016/j.neunet.2022.11.006 doi: 10.1016/j.neunet.2022.11.006
|
| [12] | Y. Kim, Convolutional neural networks for sentence classification, preprint, arXiv: 14085882. |
| [13] | D. Tang, B. Qin, T. Liu, Aspect level sentiment classification with deep memory network, preprint, arXiv: 160508900. |
| [14] | P. Lin, M. Yang, J. Lai. Deep mask memory network with semantic dependency and context moment for aspect level sentiment classification, in IJCAI, (2019), 5088–5094. DOI: 10.24963/ijcai.2019/707 |
| [15] | A. Vaswani, N. Shazeer, N. Parmar, J. Uszkoreit, L. Jones, A. N. Gomez, et al., Attention is all you need, in Advances in Neural Information Processing Systems, 30 (2017). DOI: 10.48550/arXiv.1706.03762 |
| [16] | Z.-Y. Dou, Capturing user and product information for document level sentiment analysis with deep memory network, in Proceedings of the 2017 Conference on Empirical Methods in Natural Language Processing, 2017. DOI: 10.18653/v1/D17-1054 |
| [17] |
K. Chakraborty, S. Bhattacharyya, R. Bag, A survey of sentiment analysis from social media data, IEEE Trans. Comput. Soc. Syst., 7 (2020), 450–464. DOI: 10.1109/TCSS.2019.2956957 doi: 10.1109/TCSS.2019.2956957
|
| [18] |
X. Zhu, Y. Zhu, L. Zhang, Y. Chen, A BERT-based multi-semantic learning model with aspect-aware enhancement for aspect polarity classification, Appl. Intell., 53 (2023), 4609–4623. DOI: 10.1007/s10489-022-03702-1 doi: 10.1007/s10489-022-03702-1
|
| [19] | J. Devlin, M. W. Chang, K. Lee, K. Toutanova, Bert: Pre-training of deep bidirectional transformers for language understanding, preprint, arXiv: 181004805. |
| [20] | N. Reimers, I. Gurevych, Sentence-bert: Sentence embeddings using siamese bert-networks, preprint, arXiv: 190810084. |
| [21] | L. Breiman, J. Friedman, C. J. Stone, R. A. Olshen, Classification and Regression Trees (CART), Biometrics, 1984 (1984). DOI: 10.2307/2530946 |
| [22] |
N. S. Altman, An introduction to kernel and nearest-neighbor nonparametric regression, Am. Stat., 46 (1992), 175–185. DOI: 10.1080/00031305.1992.10475879 doi: 10.1080/00031305.1992.10475879
|
| [23] | I. Rish, An empirical study of the naive Bayes classifier, in IJCAI 2001 workshop on empirical methods in artificial intelligence, (2001), 41–46. DOI: 10.1109/CSCI46756.2018.00065 |
| [24] | D. W. Hosmer Jr, S. Lemeshow, R. X. Sturdivant, Applied Logistic Regression, John Wiley & Sons, 2013. DOI: 10.1002/9781118548387 |
| [25] | C. Cortes, V. Vapnik, Support-vector networks, Mach. Learn., 20 (1995), 273–297. |
| [26] | L. Breiman, Random forests, Mach. Learn., 45 (2001), 5–32. DOI: 10.1023/A:1022627411411 |
| [27] | N. S. Joshi, S. A. Itkat, A survey on feature level sentiment analysis, Int. J. Comput. Sci. Inf. Technol., 5 (2014), 5422–5425. |
| [28] |
E. Cambria, B. White, Jumping NLP curves: A review of natural language processing research, IEEE Comput. Intell. Mag., 9 (2014), 48–57. DOI: 10.1109/MCI.2014.2307227 doi: 10.1109/MCI.2014.2307227
|
| [29] |
B. Zhang, X. Fu, C. Luo, Y. Ye, X. Li, L. Jing, Cross-domain aspect-based sentiment classification by exploiting domain-invariant semantic-primary feature, IEEE Trans. Affect. Comput., 2023 (2023), forthcoming. DOI: 10.1109/TAFFC.2023.3239540 doi: 10.1109/TAFFC.2023.3239540
|
| [30] |
H. Huang, B. Zhang, L. Jing, X. Fu, X. Chen, J. Shi, Logic tensor network with massive learned knowledge for aspect-based sentiment analysis, Knowl. Based Syst., 257 (2022), 109943. DOI: 10.1016/j.knosys.2022.109943 doi: 10.1016/j.knosys.2022.109943
|
| [31] |
X. Mei, Y. Zhou, C. Zhu, M. Wu, M. Li, S. Pan, A disentangled linguistic graph model for explainable aspect-based sentiment analysis, Knowl. Based Syst, 260 (2023), 110150. DOI: 10.1016/j.knosys.2022.110150 doi: 10.1016/j.knosys.2022.110150
|
| [32] | B. Zhang, X. Huang, Z. Huang, H. Huang, B. Zhang, X. Fu, et al., Sentiment interpretable logic tensor network for aspect-term sentiment analysis, in Proceedings of the 29th International Conference on Computational Linguistics, (2022), 6705–6714. |
| [33] | B. Xu, X. Wang, B. Yang, Z. Kang, Target embedding and position attention with lstm for aspect based sentiment analysis, in Proceedings of the 2020 5th International Conference on Mathematics and Artificial Intelligence, (2020), 93–97. DOI: 10.1145/3395260.3395280 |
| [34] | Y. Ma, H. Peng, E. Cambria, Targeted aspect-based sentiment analysis via embedding commonsense knowledge into an attentive LSTM, in Proceedings of the AAAI conference on artificial intelligence, (2018). DOI: 10.1609/aaai.v32i1.12048 |
| [35] | L. Bao, P. Lambert, T. Badia, Attention and lexicon regularized LSTM for aspect-based sentiment analysis, in Proceedings of the 57th annual meeting of the association for computational linguistics: student research workshop, (2019), 253–259. DOI: 10.18653/v1/P19-2035 |
| [36] |
Y. Xing, C. Xiao, Y. Wu, Z. Ding, A convolutional neural network for aspect-level sentiment classification, Int. J. Pattern Recognit. Artif Intell., 33 (2019), 1959046. DOI: 10.18653/v1/2021.textgraphs-1.8 doi: 10.18653/v1/2021.textgraphs-1.8
|
| [37] |
X. Wang, F. Li, Z. Zhang, G. Xu, J. Zhang, X. Sun, A unified position-aware convolutional neural network for aspect based sentiment analysis, Neurocomputing, 450 (2021), 91–103. DOI: 10.1016/j.neucom.2021.03.092 doi: 10.1016/j.neucom.2021.03.092
|
| [38] |
C. Gan, L. Wang, Z. Zhang, Z. Wang, Sparse attention based separable dilated convolutional neural network for targeted sentiment analysis, Knowl. Based Syst., 188 (2020), 104827. DOI: 10.1016/j.knosys.2019.06.035 doi: 10.1016/j.knosys.2019.06.035
|
| [39] |
N. Zhao, H. Gao, X. Wen, H. Li, Combination of convolutional neural network and gated recurrent unit for aspect-based sentiment analysis, IEEE Access, 9 (2021), 15561–15569. DOI: 10.1109/ACCESS.2021.3052937 doi: 10.1109/ACCESS.2021.3052937
|
| [40] | Y. Tay, L. A. Tuan, S. C. Hui, Dyadic memory networks for aspect-based sentiment analysis, in Proceedings of the 2017 ACM on Conference on Information and Knowledge Management, (2017), 107–116. DOI: 10.1145/3132847.3132936 |
| [41] |
Y. Chen, T. Zhuang, K. Guo, Memory network with hierarchical multi-head attention for aspect-based sentiment analysis, Appl. Intell., 51 (2021), 4287–4304. DOI: 10.1007/s10489-020-02069-5 doi: 10.1007/s10489-020-02069-5
|
| [42] |
Y. Zhang, B. Xu, T. Zhao, Convolutional multi-head self-attention on memory for aspect sentiment classification, IEEE-CAA J. Automatica Sin., 7 (2020), 1038–1044. DOI: 10.1109/JAS.2020.1003243 doi: 10.1109/JAS.2020.1003243
|
| [43] | Y. Song, J. Wang, T. Jiang, Z. Liu, Y. Rao, Attentional encoder network for targeted sentiment classification, preprint, arXiv: 190209314. |
| [44] |
H. Yang, B. Zeng, J. Yang, Y. Song, R. Xu, A multi-task learning model for chinese-oriented aspect polarity classification and aspect term extraction, Neurocomputing, 419 (2021), 344–356. DOI: 10.1016/j.neucom.2020.08.001 doi: 10.1016/j.neucom.2020.08.001
|
| [45] | A. Karimi, L. Rossi, A. Prati, Improving bert performance for aspect-based sentiment analysis, preprint, arXiv: 201011731. |
| [46] | A. Karimi, L. Rossi, A. Prati, Adversarial training for aspect-based sentiment analysis with bert, in 2020 25th International conference on pattern recognition (ICPR), (2021), 8797–8803. DOI: 10.1109/ICPR48806.2021.9412167 |
| [47] | H. Peng, Y. Ma, Y. Li, E. Cambria, Learning multi-grained aspect target sequence for Chinese sentiment analysis, Knowl. Based Syst., 148 (2018), 167–176. |
| [48] | W. Che, Y. Zhao, H. Guo, Z. Su, T. Liu, Sentence compression for aspect-based sentiment analysis, IEEE-ACM Trans. Audio Speech Lang., 23 (2015), 2111–2124. |
| [49] | L. Dong, F. Wei, C. Tan, D. Tang, M. Zhou, K. Xu, Adaptive recursive neural network for target-dependent twitter sentiment classification, in Proceedings of the 52nd annual meeting of the association for computational linguistics (volume 2: Short papers), (2014), 49–54. |
| [50] | B. Wang, W. Lu, Learning latent opinions for aspect-level sentiment classification, in Proceedings of the AAAI Conference on Artificial Intelligence, 2018. |
| [51] | H. T. Nguyen, M. Le Nguyen, Effective attention networks for aspect-level sentiment classification, in 2018 10th International Conference on Knowledge and Systems Engineering (KSE), (2018), 25–30. DOI: 10.1109/KSE.2018.8573324 |
| [52] | D. P. Kingma, J. Ba, Adam: A method for stochastic optimization, preprint, arXiv: 14126980. |
| [53] | Y. Wang, M. Huang, X. Zhu, L. Zhao, Attention-based LSTM for aspect-level sentiment classification, in Proceedings of the 2016 conference on empirical methods in natural language processing, (2016), 606–615. DOI: 10.18653/v1/D16-1058 |
| [54] | D. Ma, S. Li, X. Zhang, H. Wang, Interactive attention networks for aspect-level sentiment classification, preprint, arXiv: 170900893. |
| [55] | H. Peng, L. Xu, L. Bing, F. Huang, W. Lu, L. Si, Knowing what, how and why: A near complete solution for aspect-based sentiment analysis, in Proceedings of the AAAI conference on artificial intelligence, (2020), 8600–8607. DOI: 10.1609/aaai.v34i05.6383 |
| [56] | W. Song, Z. Wen, Z. Xiao, S. C. Park, Semantics perception and refinement network for aspect-based sentiment analysis, Knowl. Based Syst., 214 (2021), 106755. |
| [57] | L. Xu, L. Bing, W. Lu, F. Huang, Aspect sentiment classification with aspect-specific opinion spans, in Proceedings of the 2020 Conference on Empirical Methods in Natural Language Processing (EMNLP), (2020), 3561–3567. DOI: 10.18653/v1/2020.emnlp-main.288 |
| [58] |
Q. Xu, L. Zhu, T. Dai, C. Yan, Aspect-based sentiment classification with multi-attention network, Neurocomputing, 388 (2020), 135–143. DOI: 10.1016/j.neucom.2020.01.024 doi: 10.1016/j.neucom.2020.01.024
|
| [59] |
B. Huang, J. Zhang, J. Ju, R. Guo, H. Fujita, J. Liu, CRF-GCN: An effective syntactic dependency model for aspect-level sentiment analysis, Knowl. Based Syst., 260 (2023), 110125. DOI: 10.1016/j.knosys.2022.110125 doi: 10.1016/j.knosys.2022.110125
|
| [60] |
B. Huang, R. Guo, Y. Zhu, Z. Fang, G. Zeng, J. Liu, et al., Aspect-level sentiment analysis with aspect-specific context position information, Knowl. Based Syst., 243 (2022), 108473. DOI:10.1016/j.knosys.2022.108473 doi: 10.1016/j.knosys.2022.108473
|




Figures(5) / Tables(3)

Falah Mustafa Al-Saraireh. Cold-curing mixtures based on biopolymer lignin complex for casting production in single and small-series conditions[J]. AIMS Materials Science, 2023, 10(5): 876-890. doi: 10.3934/matersci.2023047







 DownLoad:
DownLoad: