




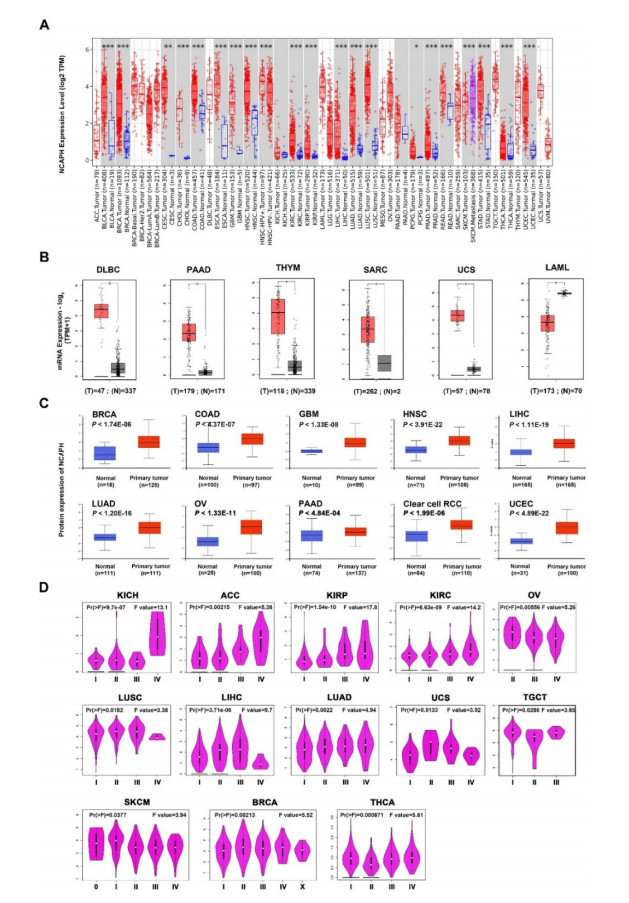
Figure 1.
Expression of NCAPH in different cancers and normal tissues. (A) TIMER2 database. (B) GEPIA2 databases. (C) CPTAC databases. (D) GEPIA2 database. (*P < 0.05, **P < 0.01, ***P < 0.001).
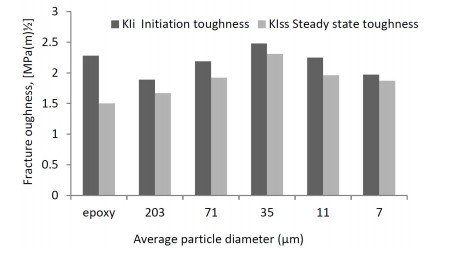
Citation: Mulugeta H. Woldemariam, Giovanni Belingardi, Ermias G. Koricho, Daniel T. Reda. Effects of nanomaterials and particles on mechanical properties and fracture toughness of composite materials: a short review[J]. AIMS Materials Science, 2019, 6(6): 1191-1212. doi: 10.3934/matersci.2019.6.1191
| [1] | Tatyana V. Polyudova, Daria V. Eroshenko, Vladimir P. Korobov . Plasma, serum, albumin, and divalent metal ions inhibit the adhesion and the biofilm formation of Cutibacterium (Propionibacterium) acnes. AIMS Microbiology, 2018, 4(1): 165-172. doi: 10.3934/microbiol.2018.1.165 |
| [2] | Meiliyana Wijaya, Ryan Halleyantoro, Jane Florida Kalumpiu . Biofilm: The invisible culprit in catheter-induced candidemia. AIMS Microbiology, 2023, 9(3): 467-485. doi: 10.3934/microbiol.2023025 |
| [3] | Joseph O. Falkinham . Mycobacterium avium complex: Adherence as a way of life. AIMS Microbiology, 2018, 4(3): 428-438. doi: 10.3934/microbiol.2018.3.428 |
| [4] | Luciana C. Gomes, Joana M. R. Moreira, José D. P. Araújo, Filipe J. Mergulhão . Surface conditioning with Escherichia coli cell wall components can reduce biofilm formation by decreasing initial adhesion. AIMS Microbiology, 2017, 3(3): 613-628. doi: 10.3934/microbiol.2017.3.613 |
| [5] | Joice Cavalcanti Lima, Lívia de Souza Ramos, Pedro Fernandes Barbosa, Iuri Casemiro Barcellos, Marta Helena Branquinha, André Luis Souza dos Santos . Biofilm production by the multidrug-resistant fungus Candida haemulonii is affected by aspartic peptidase inhibitor. AIMS Microbiology, 2025, 11(1): 228-241. doi: 10.3934/microbiol.2025012 |
| [6] | Stephen H. Kasper, Ryan Hart, Magnus Bergkvist, Rabi A. Musah, Nathaniel C. Cady . Zein nanocapsules as a tool for surface passivation, drug delivery and biofilm prevention. AIMS Microbiology, 2016, 2(4): 422-433. doi: 10.3934/microbiol.2016.4.422 |
| [7] | McKenna J. Cruikshank, Justine M. Pitzer, Kimia Ameri, Caleb V. Rother, Kathryn Cooper, Austin S. Nuxoll . Characterization of Staphylococcus lugdunensis biofilms through ethyl methanesulfonate mutagenesis. AIMS Microbiology, 2024, 10(4): 880-893. doi: 10.3934/microbiol.2024038 |
| [8] | Ariel J. Santiago, Maria L. Burgos-Garay, Leila Kartforosh, Mustafa Mazher, Rodney M. Donlan . Bacteriophage treatment of carbapenemase-producing Klebsiella pneumoniae in a multispecies biofilm: a potential biocontrol strategy for healthcare facilities. AIMS Microbiology, 2020, 6(1): 43-63. doi: 10.3934/microbiol.2020003 |
| [9] | Afraa Said Al-Adawi, Christine C. Gaylarde, Jan Sunner, Iwona B. Beech . Transfer of bacteria between stainless steel and chicken meat: A CLSM and DGGE study of biofilms. AIMS Microbiology, 2016, 2(3): 340-358. doi: 10.3934/microbiol.2016.3.340 |
| [10] | Stephen T. Abedon . Phage “delay” towards enhancing bacterial escape from biofilms: a more comprehensive way of viewing resistance to bacteriophages. AIMS Microbiology, 2017, 3(2): 186-226. doi: 10.3934/microbiol.2017.2.186 |
Cancer is considered to be the "number one killer" of human beings and is called an incurable disease [1]. Cancer has become a major enemy that endangers human health and life, and it predicts a poor prognosis of patients with cancer [2]. In recent years, relevant studies have shown that the occurrence and development of cancer are closely related to environmental factors and lifestyle changes [3]. To better understand the progression of cancer, joint efforts are needed in many aspects.
Non-chromosomal structure maintenance protein condensin complex I subunit H (non-SMC condensing I complex subunit H, NCAPH) encoded by a gene located on chromosome 2q11.2, belongs to the bar gene family and a regulatory subunit of the condensin complex [4,5,6,7]. It has been found to be associated with cell proliferation and plays a key role in mitotic chromosome structure and segregation [8,9,10]. Many studies have shown that NCAPH is involved in the evolution of various malignant tumors. Sun et al. found that NCAPH is significantly higher in cancer tissues than in normal non-cancerous tissue, and its high expression is closely correlated with the poor prognosis of patients in hepatocellular carcinoma [11]. It has been reported that high expression of NCAPH promoted the proliferation, migration, and invasion of gastric cancer cells in vitro and in vivo [12]. However, the progression of the specific mechanism of NCAPH in cancer immunology has not been well validated yet.
It is well known that tumor microenvironment (TME) is mainly composed of immune cells including CD8+T cells, CD4+T cells, regulatory T cells, tumor-associated macrophages (TAMs), tumor-associated neutrophils, and natural killer (NK) cells. They interact in the complex TME to regulate tumor development, invasion and metastasis [13,14]. In most cases, the main role of the TME is immunosuppression, which is able to inhibit the activation of lymphocytes in tumor tissue and significantly affect the antitumor effects of drugs. However, current immunotherapy works only in a subset of patients, so we need to find more effective therapeutic targets.
In this study, we accessed the TCGA and GEO databases to analyze the significance of NCAPH in cancer. Various databases such as TIMER, GSEA, Kaplan-Meier Plotter, etc. have shown that NCAPH has significant clinical correlations with the poor prognosis, DNA repair and replication, protein phosphorylation, immune cell infiltration and immune markers. These results revealed the prognostic value of NCAPH and a clear correlation between NCAPH and immune infiltration in pan-cancer.
TIMER2 database (http://timer.cistrome.org/) uses RNA-Seq expression profiling data to detect the infiltration of immune cells in tumor tissues. TIMER provides the infiltration of six types of immune cells (B cells, CD4+T cells, CD8+T cells, Neutrphils, Macrophages and Dendritic cells). We analyzed the different expression of NCAPH in tumor tissues and the correlation with immune infiltration with TIMER algorithm [15].
For tumors that are without normal tissues data, the module of GEPIA2 (http://gepia2.cancer-pku.cn/#analysis) was further adopted to analyze the box plots of NCAPH expression in the remaining cancers [16]. GEPIA2 also contains cancers of genotype-tissue expression (GTEx) database, which provides more convincing evidence. Moreover, the GEPIA2 database could also be utilized to retrieve the NCAPH expression in different pathological stages (including stage Ⅰ–Ⅳ).
The UALCAN portal (http://ualcan.path.uab.edu/analysis-prot.html) was intended to analyze protein expression by taking advantage of data from clinical proteomic tumor analysis consortium (CPTAC) [17]. We obtained available datasets of multiple tumors to explore the total protein expression of NCAPH, including its expression in liver, lung, cervical, glioma, and pancreatic cancers.
GEPIA2 database was performed to generate the survival significance map data of NCAPH more than 30 different forms of cancer, consisting of RFS (Recurrence Free Survival), DMFS (Distant Metastasis Free Survival), PPS (Post Progression Survival), PFS (Progression Free Survival), DSS (Disease Specific Survival), and FP (Full Period of Service). We also obtained the survival plots through the Kaplan-Meier plotter database, which utilized the log-rank test for hypothesis test.
The cBioPortal (http://www.cbioportal.org) web was applied to explore the mutation frequency, copy number alteration and mutation type of NCAPH [18]. We also obtained the detailed genetic alteration information in the mutations section. In addition, database searches also uncovered associations of NCAPH gene alterations with specific cancer prognoses (OS, DSS, DFS, RFS, and PFS).
PhosphoSite Plus (https://www.phosphosite.org) website supplied comprehensive information support to study on protein post-translational modifications (PTMs) [19]. Using this website, we obtained the phosphorylation features of the NCAPH protein. The diversity of phosphorylation alteration of NCAPH protein in tumor and normal tissues was further analyzed via the UALCAN portal. The expression and localization of human proteins in different tissues and organs can be retrieved through HPA (https://www.proteinatlas.org) [20]. Furthermore, HPA detected NCAPH expression in a number of different cancer types.
The TIMER1 database (https://cistrome.shinyapps.io/timer/) was originally a database used to retrieve tumor-infiltrating immune cell (TIIC) abundance in all TCGA tumors, and it was the first version of TIMER2 database [21]. Herein, the TIMER1 database was used to analyze the relevance of NCAPH and TIICs in different cancers. Then, the TIMER2 database further performed to re-evaluate the relationship among three TIICs and NCAPH expression in the overall cancer level.
The STRING (https://string-db.org/) was defined to display protein-protein interaction networks, and can be used for functional enrichment analysis [22]. Therefore, we first retrieved the top 50 proteins with potential binding to NCAPH via the site. Subsequently, we identified the top 100 targeted genes that may be related to NCAPH from the "Similar Gene Detection" module of GEPIA2. Jvenn supported intersection analysis and produce Venn diagrams [23].
We performed gene ontology (GO) enrichment and kyoto encyclopedia of genes and genomes (KEGG) pathway analysis through bioinformatics resources web server applied for annotation, visualization and integrated discovery (DAVID) [24,25]. Functional annotation charts of KEGG pathway and GO enrichment analysis which that included biological process (BP), cellular component (CC), and molecular function (MF), were plotted using bioinformatics, freely available online. (http://www.bioinformatics.com.cn) [26].
Data changes on NCAPH were obtained through the cBioPortal website, and data related to its mRNA expression were retrieved. The prognostic significance of each variable in the articles was assessed by Kaplan-Meier survival curves and compared using the log-rank test [27]. Meanwhile, Spearman's correlation test was used for correlation analysis. Unless otherwise specified (P < 0.05), the difference was statistically significant.
Difference in expression of NCAPH between human cancer and normal tissues was discovered via TIMER2 database. According to Figure 1A, NCAPH in tumor tissues was obviously higher than that of normal tissues, including bladder urothelial carcinoma (BLCA), breast invasive carcinoma (BRCA), cholangiocarcinoma (CHOL), colon adenocarcinoma (COAD), esophageal carcinoma (ESCA), glioblastoma multiforme (GBM), head and neck squamous cell carcinoma (HNSC), kidney chromophobe (KICH), kidney renal clear cell carcinoma (KIRC), kidney renal papillary cell carcinoma (KIRP), liver hepatocellular carcinoma (LIHC), lung adenocarcinoma (LUAD), lung squamous cell carcinoma (LUSC), pancreatic adenocarcinoma (PAAD), prostate adenocarcinoma (PRAD), rectum adenocarcinoma (READ), stomach adenocarcinoma (STAD), uterine corpus endometrial carcinoma (UCEC) (P < 0.05), cervical squamous cell carcinoma and endocervical adenocarcinoma (CESC), and pheochromocytoma and paraganglioma (PCPG) (P < 0.01), but it was no markedly change in thyroid carcinoma (THCA). Similarly, the GEPIA2 database was searched for NCAPH mRNA expression in pan-cancer tissues. It was up-regulated significantly in lymphoid neoplasm diffuse large B-cell lymphoma (DLBC), PAAD, thymoma (THYM), sarcoma (SARC) and uterine carcinosarcoma (UCS) compared with that of normal tissues, while down-regulated in acute myeloid leukemia (LAML) (Figure 1B, P < 0.05).
Consistent with the results, we found that the total protein level of NCAPH was markedly elevated in primary tissues than in normal tissues based on the CPTAC dataset (Figure 1C, P < 0.001). The correlation between NCAPH and the clinical stage of various tumors suggested that the expression of NCAPH was related to the clinical stage of certain cancers, consisting of KICH, adrenocortical carcinoma (ACC), KIRC, KIPR, OV, LUSC, LIHC, LUAD, UCS, TGCT, SKCM, BRCA and thyroid carcinoma (THCA). In contrast, the expression of NCAPH in clinical stage Ⅳ was lower in LUSC and LIHC than in other stages (Figure 1D, P < 0.05). These data suggested that NCAPH expression was generally overexpression in cancer tissues, and it might be involved in the malignant progression of tumors.
The prognostic significance of NCAPH expression in different tumor patients was evaluated using the TCGA and GEO datasets. It turned out that the OS of patients with NCAPH overexpression was significantly shorter than patients in the low expression group, covering ACC, KIRC, KIRP, brain lower grade glioma (LGG), LIHC, LUAD, MESO, PAAD, SARC and uveal melanoma (UVM) (P < 0.05) (Figure 2A). At the same time, NCAPH overexpression was also turned out to reduce DFS in ACC, KICH, KIRC, KIRP, LGG, LIHC, PAAD, PRAD, SARC, TGCT, THCA and UVM (P < 0.05) (Figure 2B). We further found that the overexpression of NCAPH could predict unfavorable outcome in Breast Cancer (P < 0.01), Liver Cancer (P < 0.01), Lung Cancer (P < 0.01) and Ovarian Cancer (P < 0.05) through Kaplan-Meier plotter database (P < 0.05). While the overexpression of NCAPH showed patients with gastric cancer have a longer survival period, the specific molecular mechanism needs to be further studied (P < 0.05) (Figure 3). Thus, the results indicated that overexpression of NCAPH in most tumors predicts a short survival in patients.


To further elucidate the potential molecular mechanism of NCAPH in tumorigenesis, NCAPH-related genes were screened for biological function analysis. Based on STRING, we retrieved the top 50 NCAPH-binding proteins validated by experimental data, and exhibited their PPI network (Figure 4A). Figure 4B showed that NCAPH expression was notably related to cell proliferation-related genes (BUB1, R = 0.84; CCNB2, R = 0.82; CDCA5, R = 0.82) and cell cycle regulation genes (KIF2C, R = 0.84; KIFC1, R = 0.81; TPX2, R = 0.81) of the top 100 genes (P < 0.01). We also illustrated that NCAPH was positively related to the six genes (BUB1, CCNB, CDCA, KIF2C, KIFC1 and TPX2) in most types of cancer (Figure 4C). Further searching through the Venn diagram database revealed that a total of 29 genes overlapped with each other in the two groups, they are closely related to proliferation function and cell cycle regulation function (Figure 4D).

We then performed KEGG and GO enrichment analysis by combining these two datasets, which contain a total of NCAPH-related genes. The KEGG analysis revealed that these genes were primarily concentrated in ways related to DNA replication and cell cycle signaling pathway (Figure 4E). Interestingly, it was also related to the PD-L1 checkpoint pathway and the inflammatory pathway NF-kB, suggesting that there was a certain relationship between NCAPH and the immune process. GO analysis revealed that the major BP contained cell division and DNA repair (Figure 4F). From the distribution of CC, most of the CC terms were associated with the nucleoplasm (Figure 4G). For the analysis of MF (Figure 4H), it was revealed that NCAPH was mainly related to protein binding. Consistent with our previous gene association findings, these results further suggested that NCAPH may be involved in molecular mechanisms regulating cell cycle and immune escape.
With the previous results (Figure 1), the mRNA and total protein of NCAPH were overexpressed in BRCA, COAD and other cancers. The TIMER database was further selected to investigate the potential association between the expression level of NCAPH and the level of tumor-infiltrating immune cells (TIICs) in diverse cancer types. The present database showed that NCAPH expression in THCA and KIRC was positively correlated with B cells, CD8+T cells, CD4+T cells, macrophages, neutrophils and DCs (P < 0.05). In contrast, NCAPH expression was significantly negatively correlated with macrophages (r = −0.014, P = 6.68e−01) in BRCA, CD8+T cells (r = −0.129, P = 4.56e−03) in OV, CD4+T cells (r = −0.143, P = 6.32e−02) in PAAD, and neutrophils (r = −0.122, P = 1.96e−01) in THYM (Figure 5A). Figure 5B exhibited the correlation of NCAPH with the level of immune cell infiltration in other types of cancer based on sangerbox. As shown in supplementary Figures S1 and S2, which demonstrates the correlation between NCAPH and the level of immune infiltration in a variety of cancers. Next, the potential relationship between NCAPH expression and immune cell infiltration in different TCGA tumors was considered. The results displayed that NCAPH expression and CD8+T cell infiltration showed a positive correlation in KIRP, LUAD, BRCA and THYM, while negatively correlated in HNSC (HPV-) and UCEC cancers. Consistently, we observed a positive relationship between NCAPH expression and myeloid dendritic infiltration in BRCA and THYM (Figure 6A, B, P < 0.05).


Correspondingly, there was also a clear correlation between NCAPH expression levels and immune marker genes in BRCA (Figure 6C). The results suggested that in BRCA, there was a positive association with NCAPH expression in M2 (ARG1, MRC1, both P < 0.05), TAM (CD80, CD86, both P < 0.05) and B cell (CD38, P < 0.05). In particular, the expression of NCAPH was most significantly correlated with CD80 (P = 5.67e-39, R = 0.379). Moreover, Figure S3 also showed the correlation between NCAPH and immune infiltration markers in other tumors. Therefore, NCAPH may play an immunosuppressive role in tumors through CD80 and assist tumor immune escape.
To identify the mechanism by which NCAPH impacts survival, we used cBioPortal to explore alteration frequencies, including mutation, structural variant, amplification, deep deletion, and multiple alterations, of NCAPH in different cancer types (Figure 7A). Results showed that the top five cancer types with more total mutations were UCEC (5.1%), SKCM (3.38%), STAD (3.18%), KICH (3.08%), and LUAD (2.12%). For specific alteration types, amplifications of NCAPH were enriched in UCS (3.5%), BLCA (1.7%), LUSC (1.64%), HNSC (0.96%), and LUAD (0.88%). We also found one NCAPH-deep deletion was enriched in DLBC (2.08%). At the same time, missense mutation was the commonest mutational style of NCAPH gene in TCGA tumors, and D469lfs/Rfs alteration in the Cnd2 bind domain was discovered in COAD, UCEC and STAD (Figure 7B). Based on these results, we further studied the correlation of NCAPH alteration with prognosis in the top five cancer types and found the prognostic value of NCAPH alteration. The results are summarized in Figure 7C. Altered NCAPH is significantly associated with a good prognosis in HNSC (OS (p = 0.0408), PFS (p = 0.0137), DSS (p = 0.0499)) and BLCA (DSS (p = 0.0229)). In addition, NCAPH has different protein phosphorylation sites in HNSC, and its expression in tumors was also different (P < 0.001). Higher NCAPH phosphorylation level was attended in HNSC (S59, S67, S76, S190, S222 and T38, P < 0.05). Conversely, NCAPH phosphorylated S16 was downregulated in tumor tissues, with no significant difference in phosphorylated S14 (Figure 7D).

It has been reported that NCAPH is a mitosis-associated protein that plays an important role in tumorigenesis and tumor prognosis assessment [28]. However, it was unclear whether NCAPH has a common pathway in different tumor pathogenesis. We also failed to retrieve any literature or report about NCAPH in pan-cancer. Thus, the objective was to evaluate the prognostic value and the latent biological functions of NCAPH in different types of cancer.
Our analysis demonstrated that NCAPH was highly expressed in most cancers compared with normal controls and positively correlated with the clinical stage. Interestingly, NCAPH overexpression in clinical stage Ⅳ was lower in LUSC and LIHC, which may be related to factors such as cancer type and the number of patients in the database, which further leads to the heterogeneity of NCAPH function. Likewise, the aberrant NCAPH expression usually indicates an adverse prognosis for many kinds of cancer, which may serve as a prognostic biomarker in cancer patients. Lu et al. found that NCAPH protein was highly expressed in breast cancer, and its overexpression predicts poor prognosis in patients [29]. Cui et al. demonstrated that the OS was lower in prostate cancer patients with high NCAPH expression [30]. This finding was in line with our previous analysis. Whereas, we still need to fully consider other clinical features and provide more direct evidence to confirm whether the over- expression of NCAPH was involved in the regulation of tumor malignant evolution, or was merely an outcome of anti-tumor changes in normal tissue. Together, these results strongly indicated that NCAPH may have value as a prognostic indicator for cancer.
Genetic mutations acted as a significant player in the development and growth of cancer, and they can also be targets for effective treatments [31,32]. It has been reported that the NCAPH gene with a high mutation rate in cancer, and it has an aggressive phenotype [33]. Meanwhile, our research identified that missense mutation was the most frequent mutational style of NCAPH gene in TCGA tumors. Noteworthy, NCAPH gene mutations were significantly enriched in STAD, the overexpression of NCAPH in gastric cancer predicts a longer survival of patients. Therefore, we conjecture that the mutation of NCAPH at the gene locus may be closely related to the longer survival of patients. Studies had reported that phosphorylated NCAPH in multiple tumors was well correlated with tumorigenesis [34,35,36]. The phosphorylation of NCAPH at ser988 had been found to promote the occurrence of BRCA1 [37]. Herein, our results noted that NCAPH phosphorylation (S59, S67, S76, S90, S222 and S38) is higher in HNSCs, but S16 is lower in HNSCs. Whether these post-translational modification sites have clinical significance remains unclear, and further clarification was required by subsequent molecular biology experiments.
Recently, multiple related reports have obtained that protein gene mutations and protein phosphorylation modifications also affect the level of tumor immune infiltration in different cancer types [38,39,40]. Our study elucidated the correlation between NCAPH expression and immune cell infiltration levels in various tumors, especially CD8+T cells, DCs and neutrophils. Consistent with our research, Yin et al. proposed that NCAPH overexpression is associated with poor prognosis and immune infiltration in COAD [41]. Moreover, macrophages as the first line of immune defense play an important role in every stage of tumor development and are the core regulators in the TME [42]. Interestingly, NCAPH was mainly positively correlated with M2 macrophage markers, such as CD80. Meanwhile, in our study, NCAPH expression was significantly enriched in PD-L1/NF-kB pathway, DNA replication and DNA repair pathways. The discovery that the NCAPH has plays an important role in cancer immunology and facilitates future studies with larger patient populations that could determine the feasibility of clinically integrated therapy.
In summary, NCAPH is correlated with the poor prognosis of patients, protein phosphorylation, immune cell infiltration, and immunity markers in multiple cancers. NCAPH may serve as a pan-cancer prognostic biomarker and play a key role in cancer as a new immunotherapeutic target. However, there is still a shortcoming in the molecular mechanism of NCAPH regulating tumors, and the specific cell function experiments need to be further explored. It is worth noting that although we have found NCAPH in multiple online datasets as a potential cancer biomarker and associated with poor prognosis for multiple cancer patients. But so far there is no report on the expression of NCAPH in BLCA, UCEC, GBM, CHOL, etc. This requires us to carry out more studies to improve the significance of the differential expression of NCAPH in different types of cancer in prognostic evaluation.
This research was supported by the Project from the Department of Health of Jilin Province (2020J002). The National Natural Science Foundation of China (No. 82160552).
The authors declare that there is no conflict of interest.
| [1] |
Wimmer G, Schuecker C, Pettermann HE (2009) Numerical simulation of delamination in laminated composite components-A combination of a strength criterion and fracture mechanics. Compos Part B-Eng 40: 158-165. doi: 10.1016/j.compositesb.2008.10.006

|
| [2] |
Maillet I, Michel L, Rico G, et al. (2013) A new test methodology based on structural resonance for mode I fatigue delamination growth in an unidirectional composite. Compos Struct 97: 353-362. doi: 10.1016/j.compstruct.2012.10.024
|
| [3] |
Chen J, Fox D (2012) Numerical investigation into multi-delamination failure of composite T-piece specimens under mixed mode loading using a modified cohesive model. Compos Struct 94: 2010-2016. doi: 10.1016/j.compstruct.2011.12.030
|
| [4] |
Liu HY, Wang GT, Mai YW, et al. (2011) On fracture toughness of nano-particle modified epoxy. Compos Part B-Eng 42: 2170-2175. doi: 10.1016/j.compositesb.2011.05.014
|
| [5] |
Peng L, Xu J, Zhang J, et al. (2012) Mixed mode delamination growth of multidirectional composite laminates under fatigue loading. Eng Fract Mech 96: 676-686. doi: 10.1016/j.engfracmech.2012.09.033
|
| [6] |
Albedah A, Benyahia F, Dinar H, et al. (2013) Analytical formulation of the stress intensity factor for crack emanating from central holes and repaired with bonded composite patch in aircraft structures. Compos Part B-Eng 45: 852-857. doi: 10.1016/j.compositesb.2012.08.019
|
| [7] |
Wetzel B, Rosso P, Haupert F, et al. (2006) Epoxy nanocomposites-fracture and toughening mechanisms. Eng Fract Mech 73: 2375-2398. doi: 10.1016/j.engfracmech.2006.05.018
|
| [8] |
Koricho EG, Khomenko A, Haq M, et al. (2015) Effect of hybrid (micro- and nano-) fillers on impact response of GFRP composite. Compos Struct 134: 789-798. doi: 10.1016/j.compstruct.2015.08.106
|
| [9] |
Hamitouche L, Tarfaoui M, Vautrin A (2008) An interface debonding law subject to viscous regularization for avoiding instability: application to the delamination problems. Eng Fract Mech 75: 3084-3100. doi: 10.1016/j.engfracmech.2007.12.014
|
| [10] |
Hawkins Jr DA, Haque A (2014) Fracture toughness of carbon-graphene/epoxy hybrid nanocomposites. Procedia Eng 90: 176-181. doi: 10.1016/j.proeng.2014.11.833
|
| [11] |
Tsai JL, Huang BH, Cheng YL (2011) Enhancing fracture toughness of glass/epoxy composites for wind blades using silica nanoparticles and rubber particles. Procedia Eng 14: 1982-1987. doi: 10.1016/j.proeng.2011.07.249
|
| [12] |
Tang Y, Ye L, Zhang Z, et al. (2013) Interlaminar fracture toughness and CAI strength of fibre-reinforced composites with nanoparticles-A review. Compos Sci Technol 86: 26-37. doi: 10.1016/j.compscitech.2013.06.021
|
| [13] |
Zeng Y, Liu HY, Mai YW, et al. (2012) Improving interlaminar fracture toughness of carbon fibre/epoxy laminates by incorporation of nano-particles. Compos Part B-Eng 43: 90-94. doi: 10.1016/j.compositesb.2011.04.036
|
| [14] |
Short GJ, Guild FJ, Pavier MJ (2001) The effect of delamination geometry on the compressive failure of composite laminates. Compos Sci Technol 61: 2075-2086. doi: 10.1016/S0266-3538(01)00134-8
|
| [15] |
Chandrasekaran S, Sato N, Tölle F, et al. (2014) Fracture toughness and failure mechanism of graphene based epoxy composites. Compos Sci Technol 97: 90-99. doi: 10.1016/j.compscitech.2014.03.014
|
| [16] |
Achaby ME, Ennajih H, Arrakhiz FZ, et al. (2013) Modification of montmorillonite by novel geminal benzimidazolium surfactant and its use for the preparation of polymer organoclay nanocomposites. Compos Part B-Eng 51: 310-317. doi: 10.1016/j.compositesb.2013.03.009
|
| [17] |
Valentini L, Bon SB, Lopez-Manchado MA, et al. (2016) Synergistic effect of graphene nanoplatelets and carbon black in multifunctional EPDM nanocomposites. Compos Sci Technol 128: 123-130. doi: 10.1016/j.compscitech.2016.03.024
|
| [18] |
Fu SY, Feng XQ, Lauke B, et al. (2008) Effects of particle size, particle/matrix interface adhesion and particle loading on mechanical properties of particulate-polymer composites. Compos Part B-Eng 39: 933-961. doi: 10.1016/j.compositesb.2008.01.002
|
| [19] |
Dittrich B, Wartig KA, Hofmann D, et al. (2013) Flame retardancy through carbon nanomaterials: Carbon black, multiwall nanotubes, expanded graphite, multi-layer graphene and graphene in polypropylene. Polym Degrad Stabil 98: 1495-1505. doi: 10.1016/j.polymdegradstab.2013.04.009
|
| [20] |
Hakamy A, Shaikh FUA, Low IM (2015) Effect of calcined nanoclay on microstructural and mechanical properties of chemically treated hemp fabric-reinforced cement nanocomposites. Constr Build Mater 95: 882-891. doi: 10.1016/j.conbuildmat.2015.07.145
|
| [21] |
John B, CP Nair CPR, Ninan KN (2010) Effect of nanoclay on the mechanical, dynamic mechanical and thermal properties of cyan ate ester syntactic foams. Mat Sci Eng A-Struct 527: 5435-5443. doi: 10.1016/j.msea.2010.05.016
|
| [22] |
Chang H, JiaL V, Zhang H, et al. (2017) Photo responsive colorimetric immunoassay based on chitosan modified AgI/TiO2 heterojunction for highly sensitive chloramphenicol detection. Biosens Bioelectron 87: 579-586. doi: 10.1016/j.bios.2016.09.002
|
| [23] |
Santos CSC, Gabriel B, Blanchy M, et al. (2015) Industrial applications of nanoparticles-a prospective overview. Mater Today Proc 2: 456-465. doi: 10.1016/j.matpr.2015.04.056
|
| [24] |
Liang YL, Pearson RA (2010) The toughening mechanism in hybrid epoxy-silica-rubber nanocomposites (HESRNs). Polymer 51: 4880-4890. doi: 10.1016/j.polymer.2010.08.052
|
| [25] |
Cao Q, Li J, Wang E (2019) Recent advances in the synthesis and application of copper nanomaterials based on various DNA scaffolds. Biosens Bioelectron 132: 333-342. doi: 10.1016/j.bios.2019.01.046
|
| [26] |
Liu Y, Zhou J, Shen T (2013) Effect of nano-metal particles on the fracture toughness of metal-ceramic composite. Mater Design 45: 67-71. doi: 10.1016/j.matdes.2012.08.065
|
| [27] |
Tishkevich DI, Vorobjova AI, Shimanovich DL, et al. (2019) Formation and corrosion properties of Ni-based composite material in the anodic alumina porous matrix. J Alloy Compd 804: 139-146. doi: 10.1016/j.jallcom.2019.07.001
|
| [28] |
Tishkevich DI, Grabchikov SS, Lastovskii SB, et al. (2019) Function composites materials for shielding applications: correlation between phase separation and attenuation properties. J Alloy Compd 771: 238-245. doi: 10.1016/j.jallcom.2018.08.209
|
| [29] |
Tishkevich DI, Korolkov IV, Kozlovskiy AL, et al. (2019) Immobilization of boron-rich compound on Fe3O4 nanoparticles: stability and cytotoxicity. J Alloy Compd 797: 573-581. doi: 10.1016/j.jallcom.2019.05.075
|
| [30] |
Zappalorto M, Salviato M, Quaresimin M (2013) Mixed mode (I + II) fracture toughness of polymer nanoclay nanocomposites. Eng Fract Mech 111: 50-64. doi: 10.1016/j.engfracmech.2013.09.006
|
| [31] |
Chan M, Lau K, Wong T, et al. (2011) Mechanism of reinforcement in a nanoclay/polymer composite. Compos Part B-Eng 42: 1708-1712. doi: 10.1016/j.compositesb.2011.03.011
|
| [32] | Binu PP, George KE, Vinodkumar MN (2016) Effect of nanoclay, Cloisite15A on the mechanical properties and thermal behavior of glass fiber reinforced polyester. Procedia Manuf 25: 846-853. |
| [33] |
Domun N, Hadavinia H, Zhang T, et al. (2015) Improving the fracture toughness and the strength of epoxy using nanomaterials-a review of the current status. Nanoscale 7: 10294-10329. doi: 10.1039/C5NR01354B
|
| [34] |
Asadi J, Ebrahimi NG, Razzaghi-Kashani M (2015) Self-healing property of epoxy/nanoclay nanocomposite using poly(ethylene-co-methacrylic acid) agent. Compos Part A-Appl S 68: 56-61. doi: 10.1016/j.compositesa.2014.09.017
|
| [35] |
Quang TN, Donald GB (2007) An improved technique for exfoliating and dispersing nanoclay particles into polymer matrices using supercritical carbon dioxide. Polymer 48: 6923-6933. doi: 10.1016/j.polymer.2007.09.015
|
| [36] |
El-Sheikhy R, Al-Shamrani M (2017) Interfacial bond assessment of clay-polyolefin nanocomposites CPNC on view of mechanical and fracture properties. Adv Powder Technol 28: 983-992. doi: 10.1016/j.apt.2017.01.002
|
| [37] |
Azeez AA, Rhee KY, SooJin Park SJ, et al. (2013) Epoxy clay nanocomposites-processing, properties and applications: A review. Compos Part B-Eng 45: 308-320. doi: 10.1016/j.compositesb.2012.04.012
|
| [38] |
Assaedi H, Shaikh FUA, Low IM (2016) Effect of nanoclayon mechanical and thermal properties of geopolymer. J Asian Ceram Soc 4: 19-28. doi: 10.1016/j.jascer.2015.10.004
|
| [39] |
Eesaee M, Shojaei A (2014) Effect of nanoclays on the mechanical properties and durability of novolac phenolic resin/woven glass fiber composite at various chemical environments. Compos Part A-Appl S 63: 149-158. doi: 10.1016/j.compositesa.2014.04.008
|
| [40] |
Avila AF, David Morais DTS (2009) Modeling nanoclay effects into laminates failure strength and porosity. Compos Struct 87: 55-62. doi: 10.1016/j.compstruct.2007.12.009
|
| [41] | Maharsia RR, Jerro HD (2007) Enhancing tensile strength and toughness in syntactic foams through nanoclay reinforcement. Mat Sci Eng A-Struct 454: 416-422. |
| [42] | Withers GJ, Yu Y, Khabashesku VN, Cercone L, et al. (2015) Improved mechanical properties of an epoxy glass-fiber composite reinforced with surface organo modified nanoclays. Compos Part B-Eng 72: 175-182. |
| [43] | Sharma B, Mahajan S, Chhibber R, et al. (2012) Glass fiber reinforced polymer-clay nanocomposites: processing, structure and hygrothermal effects on mechanical properties. Procedia Manuf 4: 39-46. |
| [44] |
Wang L, Wang K, Chen L, et al. (2006) Preparation, morphology and thermal/mechanical properties of epoxy/nanoclay composite. Compos Part A-Appl S 37: 1890-1896. doi: 10.1016/j.compositesa.2005.12.020
|
| [45] |
Ravandi M, Teo WS, Tran LQN, et al. (2016) The effects of through-the-thickness stitching on the Mode I interlaminar fracture toughness of flax/epoxy composite laminates. Mater Design 109: 659-669. doi: 10.1016/j.matdes.2016.07.093
|
| [46] |
Giannopoulos GI (2019) Linking MD and FEM to predict the mechanical behaviour of fullerene reinforced nylon-12. Compos Part B-Eng 161: 455-463. doi: 10.1016/j.compositesb.2018.12.110
|
| [47] | Giannopoulos GI (2019) Introducing bone-shaped carbon nanotubes to reinforce polymer nanocomposites: A molecular dynamics investigation. Mater Today Chem 20: 100570. |
| [48] | Giannopoulos GI, Georgantzinos SK, Katsareas DE, et al. (2010) Numerical prediction of young's and shear moduli of carbon nanotube composites incorporating nanoscale and interfacial effects. CMES-Comp Model Eng 56: 231-247. |
| [49] |
Shuvo SN, Shorowordi KMK (2015) Processing and mechanical mharacterization of graded and non-graded nanoclay composites. Procedia Eng 105: 928-932. doi: 10.1016/j.proeng.2015.05.117
|
| [50] |
Ghadami F, Dadfar MR, Kazazi M (2016) Hot-cured epoxy-nanoparticulate-filled nanocomposites: fracture toughness behavior. Eng Fract Mech 162: 193-200. doi: 10.1016/j.engfracmech.2016.05.016
|
| [51] |
Phong NT, Gabr MH, Okubo K, et al. (2013) Improvement in the mechanical performances of carbon fiber/epoxy composite with addition of nano-(Polyvinyl alcohol) fibers. Compos Struct 99: 380-387. doi: 10.1016/j.compstruct.2012.12.018
|
| [52] |
Gantenbein D, Schoelkopf J, Peter G, et al. (2011) Determining the size distribution-defined aspect ratio of platy particles. Appl Clay Sci 53: 544-552. doi: 10.1016/j.clay.2011.04.020
|
| [53] |
Iman M, Maji TK (2012) Effect of cross linker and nanoclay on starch and jute fabric based green nanocomposites. Carbohyd Polym 89: 290-297. doi: 10.1016/j.carbpol.2012.03.012
|
| [54] |
Matusik J, Stodolak E, Bahranowski K (2011) Synthesis of polylactide/clay composites using structurally different kaolinites and kaolinite nanotubes. Appl Clay Sci 51: 102-109. doi: 10.1016/j.clay.2010.11.010
|
| [55] |
Awad WH, Beyer G, Benderly D, et al. (2009) Material properties of nanoclay PVC composites. Polymer 50: 1857-1867. doi: 10.1016/j.polymer.2009.02.007
|
| [56] | Kitey R, Tippur HV (2005) Role of particle size and filler-matrix adhesion on dynamic fracture of glass-filled epoxy. I. Macro measurements. Acta Mater 53: 1153-1165. |
| [57] |
Zhao Y, Chen Z, Liu Y, et al. (2013) Simultaneously enhanced cryogenic tensile strength and fracture toughness of epoxy resins by carboxylic nit rile-butadiene nanorubber. Compos Part A-Appl S 55: 178-187. doi: 10.1016/j.compositesa.2013.09.005
|
| [58] |
Sun XC, Wisnom MR, Hallett SR (2016) Interaction of inter- and intralaminar damage in scaled quasi-static indentation tests: part 2-Numerical simulation. Compos Struct 136: 727-742. doi: 10.1016/j.compstruct.2015.09.062
|
| [59] |
Wood MDK, Sun X, Tong L, et al. (2007) The effect of stitch distribution on Mode I delamination toughness of stitched laminated composites-experimental results and FEA simulation. Compos Sci Technol 67: 1058-1072. doi: 10.1016/j.compscitech.2006.06.002
|
| [60] |
Mouritza AP, Leongb KH, Herszberg I (1997) A review of the effect of stitching on the in-plane mechanical properties of fibre-reinforced polymer composites. Compos Part A-Appl S 28: 979-991. doi: 10.1016/S1359-835X(97)00057-2
|
| [61] | Dransfield KA, Jainb LK, Mai Y (1998) On the effects of stitching in CFRPs-I. mode I delamination toughness. Compos Sci Technol 58: 815-827. |
| [62] | Jain LK, Dransfieldb KA, Ma Y (1998) On the effects of stitching in CFRPs-II. mode II delamination toughness. Compos Sci Technol 58: 829-837. |
| [63] |
Sun X, Tong L, Wood MDK, Mai Y (2004) Effect of stitch distribution on mode I delamination toughness of laminated DCB specimens. Compos Sci Technol 64: 967-981. doi: 10.1016/j.compscitech.2003.07.004
|
| [64] |
Kim BC, Park SW, Lee DG (2008) Fracture toughness of the nano-particle reinforced epoxy composite. Compos Struct 86: 69-77. doi: 10.1016/j.compstruct.2008.03.005
|
| [65] |
Wang M, Ma L, Shi L, et al. (2019) Chemical grafting of nano-SiO2 onto graphene oxide via thiol-ene click chemistry and its effect on the interfacial and mechanical properties of GO/epoxy composites. Compos Sci Technol 182: 107751. doi: 10.1016/j.compscitech.2019.107751
|
| [66] |
Kelkar AD, Mohan R, Bolick R, et al. (2010) Effect of nanoparticles and nanofibers on Mode I fracture toughness of fiber glass reinforced polymeric matrix composites. Mat Sci Eng B-Solid 168: 85-89. doi: 10.1016/j.mseb.2010.01.015
|
| [67] |
Raja RS, Manisekar K (2016) Experimental and statistical analysis on mechanical properties of nanoflyash impregnated GFRP composites using central composite design method. Mater Design 89: 884-892. doi: 10.1016/j.matdes.2015.10.043
|
| [68] |
Moustafa H, Darwish NA (2015) Effect of different types and loadings of modified nanoclay on mechanical properties and adhesion strength of EPDM-g-MAH/nylon 66 systems. Int J Adhes Adhes 61: 15-22. doi: 10.1016/j.ijadhadh.2015.05.002
|
| [69] |
Sellappan P, Guin J, Rocherulle J, et al. (2013) Influence of diamond particles content on the critical load for crack initiation and fracture toughness of SiOC glass-diamond composites. J Eur Ceram Soc 33: 847-858. doi: 10.1016/j.jeurceramsoc.2012.10.012
|
| [70] |
Quaresimin M, Salviato M, Zappalorto M (2012) Fracture and interlaminar properties of clay-modified epoxies and their glass reinforced laminates. Eng Fract Mech 81: 80-93. doi: 10.1016/j.engfracmech.2011.10.004
|
| [71] |
Zeinedini A, Shokrieh MM, Ebrahimi A (2018) The effect of agglomeration on the fracture toughness of CNTs-reinforced nanocomposites. Theor Appl Fract Mec 94: 84-94. doi: 10.1016/j.tafmec.2018.01.009
|
| [72] |
Maghsoudlou MA, Isfahan RB, Saber-Samandari S, et al. (2019) Effect of interphase, curvature and agglomeration of SWCNTs on mechanical properties of polymer-based nanocomposites: Experimental and numerical investigations. Compos Part B-Eng 175: 107119. doi: 10.1016/j.compositesb.2019.107119
|
| [73] |
Zare Y, Rhee KP, Hui D (2017) Influences of nanoparticles aggregation/agglomeration on the interfacial/interphase and tensile properties of nanocomposites. Compos Part B-Eng 122: 41-46 doi: 10.1016/j.compositesb.2017.04.008
|
| [74] |
Rafiq A, Merah N, Boukhili R, et al. (2017) Impact resistance of hybrid glass fiber reinforced epoxy/nanoclay composite. Polym Test 57: 1-11. doi: 10.1016/j.polymertesting.2016.11.005
|
| [75] |
Domun N, Kaboglu C, Keith R (2019) Ballistic impact behaviour of glass fibre reinforced polymer composite with 1D/2D nanomodified epoxy matrices. Compos Part B-Eng 167: 497-506. doi: 10.1016/j.compositesb.2019.03.024
|
| [76] | Dorigato A, Morandi S, Pegoretti A (12) Effect of nanoclay addition on the fiber/matrix adhesion in epoxy/glass composites. J Compos Mater 46:1439-1451. |
| [77] |
Gelineau P Weigand S, Cauvin L, et al (2018) Compatibility effects of modified montmorillonite on elastic and viscoelastic properties of nano-reinforced poly(lactic acid): experimental and modeling study. Polym Test 70: 441-448. doi: 10.1016/j.polymertesting.2018.06.020
|
| [78] |
Mu X, Zhan J, Wang J, et al. (2019) A novel and efficient strategy to exfoliation of covalent organic frameworks and a significant advantage of covalent organic frameworks nanosheets as polymer nano-enhancer: high interface compatibility. J Colloid Interf Sci 539: 609-618. doi: 10.1016/j.jcis.2018.12.103
|
| [79] |
Bandyopadhyay J, Ray SS, Salehiyan R, et al. (2017) Effect of the mode of nanoclay inclusion on morphology development and rheological properties of nylon6/ethylevinyl-alcohol blend composites. Polymer 126: 96-108. doi: 10.1016/j.polymer.2017.08.032
|
| [80] | Vo VS, Nguyen V, Mahouche-Chergui S, et al. (2018) Estimation of effective elastic properties of polymer/clay nanocomposites: A parametric study. Compos Part B-Eng 152: 139-150. |
Figures(4) / Tables(3)

Mulugeta H. Woldemariam, Giovanni Belingardi, Ermias G. Koricho, Daniel T. Reda. Effects of nanomaterials and particles on mechanical properties and fracture toughness of composite materials: a short review[J]. AIMS Materials Science, 2019, 6(6): 1191-1212. doi: 10.3934/matersci.2019.6.1191

 DownLoad:
DownLoad:
