Citation: S. Pavan Kumar Naik, P. Missak Swarup Raju. Microstructural and magnetic properties of YBCO nanorods: synthesized by template growth method[J]. AIMS Materials Science, 2016, 3(3): 916-926. doi: 10.3934/matersci.2016.3.916

| [1] | Rao CNR, Kulkarni GU, Thomas PJ, et al. (2002) Size-Dependent Chemistry: Properties of Nanocrystals. Chem A Eur J 8: 28. |
| [2] | Ethayaraj M, Bandyopadhyaya R (2007) Mechanism and Modeling of Nanorod Formation from Nanodots. Langmuir 23: 6418. |
| [3] | Tachiki M, Yamashita T (1997) Sendai: Proc. 1st RIEC International Symposium on Intrinsic Josephson Effects and THz Plasma Oscillations in High-temperature Superconductors. |
| [4] | Bergeal N, Lesueur J, Sirena M, et al. (2007) Using ion irradiation to make high-Tc Josephson junctions. J Appl Phys 102: 083903. |
| [5] | Müller P, Festkörperprobleme À (1995) Adv Solid State Phys 34: 1. |
| [6] | Kleiner R, Müller P (1996) Critical Currents and Devices (New Age Int, India) 172. |
| [7] | Nagao M, Sato M, Maeda H, et al. (2003) Superconducting properties of single-crystal whiskers of(Y0.86Ca0.14)Ba2Cu3Ox (Y0.86Ca0.14)Ba2Cu3Oxgrown from precursors containing calcium and tellurium. Appl Phys Lett 82: 1899–1901. |
| [8] |
Yu AK, Golubev DS, Zaikin AD (2008) Superconductivity in one dimension Physics. Reports 464: 1-70. doi: 10.1016/j.physrep.2008.04.009

|
| [9] | Ginzburg V (1957) Ferromagnetic Superconductors. J Exp Theor Phys 4: 153. |
| [10] |
Saxena SS, Agarwal P, Ahilan K, et al. (2000) Superconductivity on the border of itinerant-electron ferromagnetism in UGe2, Superconductivity on the border of itinerant-electron ferromagnetism in UGe2. Nature 406: 587. doi: 10.1038/35020500
|
| [11] |
Gasmia M, Khene S, Fillion G (2013) J Coexistence of superconductivity and ferromagnetism in nanosized YBCO powders. J Phys Chem Solids 74: 1414. doi: 10.1016/j.jpcs.2013.04.025
|
| [12] |
Sundaresan A, Rao CNR (2009) Ferromagnetism as a universal feature of inorganic nanoparticles. Nano Today 4: 96. doi: 10.1016/j.nantod.2008.10.002
|
| [13] |
Shipra A, Gomathi A, Sundaresan A, et al. (2007) Room-temperature ferromagnetism in nanoparticles of superconducting materials. Solid State Commun 142: 685. doi: 10.1016/j.ssc.2007.04.041
|
| [14] |
Sundaresan A, Rao CNR (2009) Implications and consequences of ferromagnetism universally exhibited by inorganic nanoparticles. Solid State Commun 149: 1197. doi: 10.1016/j.ssc.2009.04.028
|
| [15] | Sundaresan A, Bhargavi R, Rangarajan N, et al. (2006) Ferromagnetism as a universal feature of nanoparticles of the otherwise nonmagnetic oxides. Phys Rev B 74: 161306. |
| [16] |
Elfimov IS, Yunoki S, Sawatzky GA (2002) Possible Path to a New Class of Ferromagnetic and Half-Metallic Ferromagnetic Materials. Phys Rev Lett 89: 216403. doi: 10.1103/PhysRevLett.89.216403
|
| [17] |
Zywietz A, Furthmuller J, Bechstedt F (2000) Spin state of vacancies: From magnetic Jahn-Teller distortions to multiplets. Phys Rev B 62: 6854. doi: 10.1103/PhysRevB.62.6854
|
| [18] | Hasanain SK, Akhtar N, Mumtaz A (2010) Particle size dependence of the superconductivity and ferromagnetism in YBCO nanoparticles. J Nanoparticle Res 13: 1953. |
| [19] | Alikhanzadeh Arani S, Salavati Niasari M (2011) Chemical Synthesis of Sub-Micrometer- to Nanometer-Sized of Antiferromagnetic Sr2CuO3 Ceramic. Proceedings of the International Conference Nanomaterials: Applications and Properties. 1: 1. |
| [20] |
Arabi H, Jamshidi S, Komeili M, et al. (2013) Coexistence of Superconductivity and Ferromagnetic Phases in YBa2Cu3O7−δNanoparticles. J Supercond Nov Magn 26: 2069. doi: 10.1007/s10948-012-1859-8
|
| [21] | Zhu Z, Gao D, Dong C, et al. (2012) Coexistence of ferromagnetism and superconductivity in YBCOnanoparticles. Phys Chem Chem Phys 14: 3859. |
| [22] | Zhang YF, Tang YH, Duan XF, et al. (2000) Yttrium–barium–copper–oxygen nanorods synthesized by laser ablation. Chem Phys Lett 323: 180. |
| [23] | Zuo JM, Mabon JC (2004) Web-based Electron Microscopy Application Software: Web-EMAPS. Microsc Microanal 10. Available from: http://emaps.mrl.uiuc.edu/. |
| [24] | Martirosyan KS, Galstyan E, Xue YY, et al. (2008) The fabrication of YBCO superconductor polycrystalline powder by CCSO. Supercond Sci Technol 21: 065008. |
| [25] | Pathak LC, Mishra SK (2005) A review on the synthesis of Y–Ba–Cu-oxide powder. Supercond Sci Technol 18: R67. |
| [26] |
Cava RJ, Batlogg B, Van Dover RB, et al. (1987) Bulk superconductivity at 91 K in single-phase oxygen-deficient perovskiteBa2YCu3O9−δ. Phys Rev Lett 58: 1676. doi: 10.1103/PhysRevLett.58.1676
|
| [27] | Bgreid TL, Fossheim K (1988) Evidence for Glasslike Dynamic Behaviour in YBa2Cu3O7-x (YBCO) Superconductor. Europhys Lett 6: 81. |
| [28] | Mook HA, Dai P, Dogan F (2002) Charge and Spin Structure inYBa2Cu3O6.35. Phys Rev Lett 88: 097004. |
| [29] | Mamalis AG, Petrov MI, Balaev DA, et al. (2001) A dc superconducting fault current limiter using die-pressed YBa2Cu3O7ceramic. Supercond Sci Technol 14: 413. |
| [30] |
Maki K (1968) The Critical Fluctuation of the Order Parameter in Type-II Superconductors. Prog Theor Phys 39: 897. doi: 10.1143/PTP.39.897
|
| [31] | Little WA (1967) Decay of Persistent Currents in Small Superconductors. Phys Rev 156: 396. |
| [32] | Zhang W-H, Sun Y, Zhang J-S, et al. (2014) Direct Observation of High-Temperature Superconductivity in One-Unit-Cell FeSe Films. Chin Phys Lett 31: 017401. |
| [33] |
Giordano N (1988) Evidence for Macroscopic Quantum Tunneling in One-Dimensional Superconductors. Phys Rev Lett 61: 2137. doi: 10.1103/PhysRevLett.61.2137
|
| [34] | Zgirski M, Riikonen KP, Tuboltsev V, et al. (2005) Size Dependent Breakdown of Superconductivity in Ultranarrow Nanowires. Nano Lett 5: 1029. |
| [35] |
Zgirski M, Riikonen KP, Tuboltsev V, et al. (2008) Quantum fluctuations in ultranarrow superconducting aluminum nanowires. Phys Rev B 77: 054508. doi: 10.1103/PhysRevB.77.054508
|
| [36] |
Altomare F, Chang AM, Melloch MR, et al. (2006) Evidence for Macroscopic Quantum Tunneling of Phase Slips in Long One-Dimensional Superconducting Al Wires. Phys Rev Lett 97: 017001. doi: 10.1103/PhysRevLett.97.017001
|
| [37] | Tian M, Kumar N, Xu S, et al. (2005) Suppression of Superconductivity in Zinc Nanowires by Bulk Superconductors. Phys Rev Lett 95: 076802. |
| [38] | Tian M, Wang J, Zhang Q, et al. (2009) Superconductivity and Quantum Oscillations in Crystalline Bi Nanowire. Nano Lett 9: 3196. |
| [39] | Wang J, Sun Y, Tian M, et al. (2012) Superconductivity in single crystalline Pb nanowires contacted by normal metal electrodes. Phys Rev B 86: 035439. |
| [40] | Bezryadin A, Lau CN, Tinkham M (2009) Quantum suppression of superconductivity in ultrathin nanowires. Nature 404: 971. |
| [41] | Bollinger AT, Reached A, Bezryadin A (2006) Dichotomy in short superconducting nanowires: Thermal phase slippage vs. Coulomb blockade. Europhys Lett 76: 505. |
| [42] |
Sharifi F, Herzog AV, Dynes RC (1993) Crossover from two to one dimension inin situgrown wires of Pb. Phys Rev Lett 71: 428. doi: 10.1103/PhysRevLett.71.428
|
| [43] | Rogachev A, Bezryadin A (2003) Superconducting properties of polycrystalline Nb nanowires templated by carbon nanotubes. Appl Phys Lett 83: 512. |
| [44] | Rogachev A, Bollinger AT, Bezryadin A (2005) Influence of High Magnetic Fields on the Superconducting Transition of One-Dimensional Nb and MoGe Nanowires. Phys Rev Lett 94: 017004. |
| [45] |
Tian M, Wang J, Kurtz J S, et al. (2005) Dissipation in quasi-one-dimensional superconducting single-crystalSnnanowires. Phys Rev B 71: 104521. doi: 10.1103/PhysRevB.71.104521
|
| [46] |
Michotte S, Piraux L, Boyer F, et al. (2004) Development of phase-slip centers in superconductingSnSnnanowires. Appl Phys Lett 85: 3175. doi: 10.1063/1.1804608
|
| [47] |
Piraux L, Encinas A, Vila L, et al. (2005) Magnetic and Superconducting Nanowires. Nanosci Nano Technol 5: 372. doi: 10.1166/jnn.2005.062
|
| [48] | Lukens JE, Warburton RJ, Webb WW (1970) Onset of Quantized Thermal Fluctuations in One-Dimensional Superconductors. Phys Rev Lett 25: 1180. |
| [49] |
Newbower RS, Beasley MR, Tinkham M (1972) Fluctuation Effects on the Superconducting Transition of Tin Whisker Crystals. Phys Rev B 5: 864. doi: 10.1103/PhysRevB.5.864
|
| [50] |
Muralt P, Pohl DW (1986) Scanning tunneling potentiometry. Appl Phys Lett 48: 514. doi: 10.1063/1.96491
|
| [51] |
Soniera JE, Kaisera CV, Pacradounia V, et al. (2010) Direct search for a ferromagnetic phase in a heavily overdoped nonsuperconducting copper oxide. P Natl Acad Sci 107: 17131. doi: 10.1073/pnas.1007079107
|
| [52] | Barbiellini B, Jarlborg T (2008) Evidence for Macroscopic Quantum Tunneling of Phase Slips in Long One-Dimensional Superconducting Al Wires. Phys Rev Lett 101: 157002. |

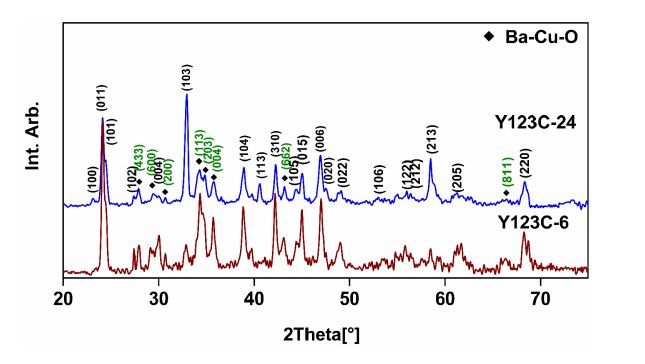
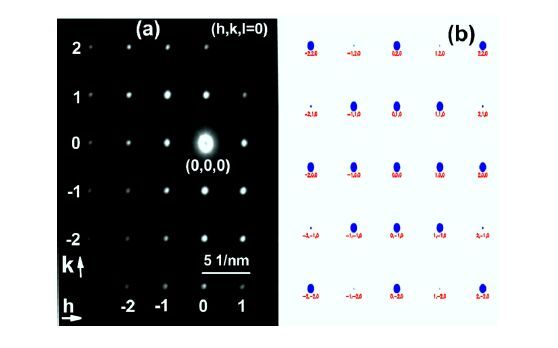
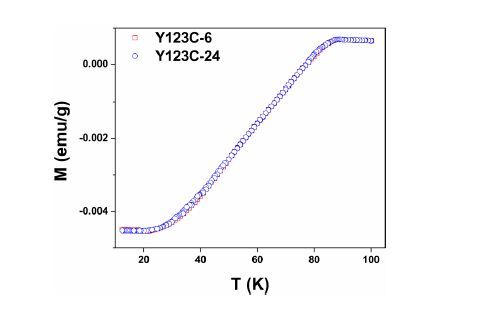
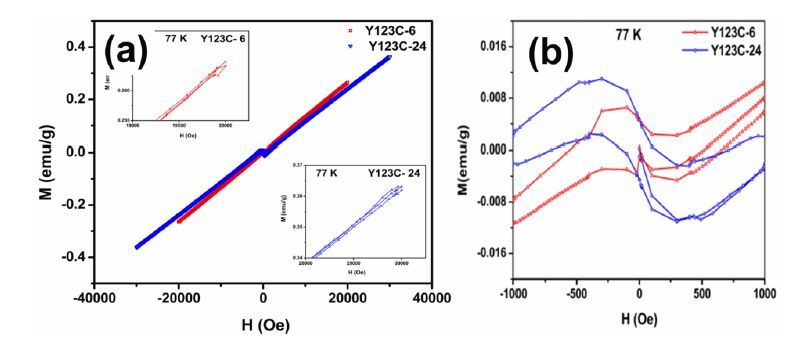
Figures(6)

S. Pavan Kumar Naik, P. Missak Swarup Raju. Microstructural and magnetic properties of YBCO nanorods: synthesized by template growth method[J]. AIMS Materials Science, 2016, 3(3): 916-926. doi: 10.3934/matersci.2016.3.916







 DownLoad:
DownLoad: