Citation: Rudolf Kawalla, Anett Stöcker, Ulrich Prahl, Xuefei Wei, Jens Dierdorf, Gerhard Hirt, Martin Heller, Stefan Roggenbuck, Sandra Korte-Kerzel, Hannes Alois Weiss, Philipp Tröber, Lucas Böhm, Wolfram Volk, Nora Leuning, Kay Hameyer. Low-loss FeSi sheet for energy-efficient electrical drives[J]. AIMS Materials Science, 2018, 5(6): 1184-1198. doi: 10.3934/matersci.2018.6.1184

| [1] |
Cui S, Jung IH (2017) Critical reassessment of the Fe-Si system. Calphad 56: 108–125. doi: 10.1016/j.calphad.2016.11.003

|
| [2] |
Schmidtchen M, Kawalla R (2016) Fast numerical simulation of symmetric flat rolling processes for inhomogeneous materials using a layer model-Part I: Basic theory. Steel Res Int 87: 1065–1081. doi: 10.1002/srin.201600047
|
| [3] | Stöcker A, Schmidtchen M, Kawalla R (2017) Hot rolling simulation for non-oriented electrical steel. AIP Conf Proc 1896: 190021. |
| [4] | Wei X, Hojda S, Dierdorf J, et al. (2016) Crystal plasticity finite element analysis of texture evolution during cold rolling of a non-oriented electrical steel. 10th International Rolling Conference and the 7th European Rolling Conference, 494–504. |
| [5] | Wei X, Hojda S, Dierdorf J, et al. (2017) Model for texture evolution in cold rolling of 2.4 wt.-% Si non-oriented electrical steel. AIP Conf Proc 1896: 170005. |
| [6] | Roters F, Diehl M (2018) "DAMASK", Max-Planck-Institut für Eisenforschung. Available from: https://damask.mpie.de/Home/WebHome. |
| [7] |
Mießen C, Liesenjohann M, Barrales-Mora LA, et al. (2015) An advanced level set approach to grain growth-Accounting for grain boundary anisotropy and finite triple junction mobility. Acta Mater 99: 39–48. doi: 10.1016/j.actamat.2015.07.040
|
| [8] | IMM Microstructure Generator, 2017. Available from: https://github.com/GraGLeS/IMM_MicrostructureGenerator. |
| [9] | GraGLeS2D-Grain Growth Level Set 2dim, 2017. Available from: https://github.com/GraGLeS/GraGLeS2D. |
| [10] |
Leuning N, Steentjes S, Stöcker A, et al. (2018) Impact of the interaction of material production and mechanical processing on the magnetic properties of non-oriented electrical steel. AIP Adv 8: 047601. doi: 10.1063/1.4994143
|
| [11] | Kestens L, Jacobs S (2008) Texture control during the manufacturing of nonoriented electrical steels. Texture Stress Microstruct 2018: 173083. |
| [12] |
Weiss H, Leuning N, Steentjes S, et al. (2017) Influence of shear cutting parameters on the electromagnetic properties of non-oriented electrical steel sheets. J Magn Magn Mater 421: 250–259. doi: 10.1016/j.jmmm.2016.08.002
|
| [13] | Weiss H, Trober P, Golle R, et al. (2017) Loss reduction due to blanking parameter optimization for different non-grain oriented electrical steel grades. IEEE International Electric Machines and Drives Conference (IEMDC). |
| [14] |
Leuning N, Steentjes S, Schulte M, et al. (2016) Effect of elastic and plastic tensile mechanical loading on the magnetic properties of NGO electrical steel. J Magn Magn Mater 417: 42–48. doi: 10.1016/j.jmmm.2016.05.049
|
| [15] |
Moses AJ (2012) Energy efficient electrical steels: Magnetic performance prediction and optimization. Scripta Mater 67: 560–565. doi: 10.1016/j.scriptamat.2012.02.027
|
| [16] | Steentjes S, Leuning N, Dierdorf J, et al. (2016) Effect of the interdependence of cold rolling strategie and subsequent punching on magnetic properties of NO steel sheets. IEEE T Magn 52: 1–4. |
| [17] | Lee K, Park S, Huh M, et al. (2014) Effect of texture and grain size on magnetic flux density and core loss in non-oriented electrical steel containing 3.15% Si. J Magn Magn Mater 354: 324–332. |
| [18] |
Leuning N, Steentjes S, Hameyer K (2019) Effect of grain size and magnetic texture on iron-loss components in NO electrical steel at different frequencies. J Magn Magn Mater 469: 373–382. doi: 10.1016/j.jmmm.2018.07.073
|
| [19] |
Pfeifer F, Kunz W (1977) Bedeutung von Kornstruktur und Fremdkörpereinschlüssen für die Magnetisierungseigenschaften hochpermeabler Ni-Fe-legierungen. J Magn Magn Mater 4: 214–219. doi: 10.1016/0304-8853(77)90038-5
|
| [20] |
Bertotti G (1988) General properties of power losses in soft ferromagnetic materials. IEEE T Magn 24: 621–630. doi: 10.1109/20.43994
|
| [21] | Lordache VE, Hug E, Buiron N (2003) Magnetic behaviour versus tensile deformation mechanisms in a non-oriented Fe–(3 wt.%)Si steel. Mat Sci Eng A-Struct 359: 62–74. |
| [22] |
Campos M, Teixeira JC, Landgraf F (2006) The optimum grain size for minimizing energy losses in iron. J Magn Magn Mater 301: 94–99. doi: 10.1016/j.jmmm.2005.06.014
|
| [23] |
Barros J, Schneider J, Verbeken K, et al. (2008) On the correlation between microstructure and magnetic losses in electrical steel. J Magn Magn Mater 320: 2490–2493. doi: 10.1016/j.jmmm.2008.04.056
|
| [24] |
Landgraf F, da Silveira J, Rodrigues Jr. D (2011) Determining the effect of grain size and maximum induction upon coercive field of electrical steels. J Magn Magn Mater 323: 2335–2339. doi: 10.1016/j.jmmm.2011.03.034
|
| [25] | Leuning N, Steentjes S, Hameyer K (2017) Effect of magnetic anisotropy on Villari Effect in non-oriented FeSi electrical steel. Int J Appl Electrom 55: 23–31. |
| [26] | Bertotti G (1998) Hysteresis in Magnetism: For Physicists, Materials Scientists, and Engineers, Academic Press, San Diego. |
| [27] |
Eggers D, Steentjes S, Hameyer K (2012) Advanced iron-loss estimation for nonlinear material behavior. IEEE T Magn 48: 3021–3024. doi: 10.1109/TMAG.2012.2208944
|
| [28] | Steentjes S, Leßmann M, Hameyer K (2013) Semi-physical parameter identification for an iron-loss formula allowing loss-separation. J Appl Phys 113: 17A319. |
| [29] |
Leuning N, Steentjes S, Hameyer K (2017) On the homogeneity and isotropy of non-grain-oriented electrical steel sheets for the modeling of basic magnetic properties from microstructure and texture. IEEE T Magn 53: 1–5. doi: 10.1109/TMAG.2018.2792846
|
| [30] | Ruf A, Steentjes S, von Pfingsten G, et al. (2016) Requirements on soft magnetic materials for electric traction motors. Proceedings: 7th International Conference on Magnetism and Metallurgy: WMM'16, Rome, Italy. |
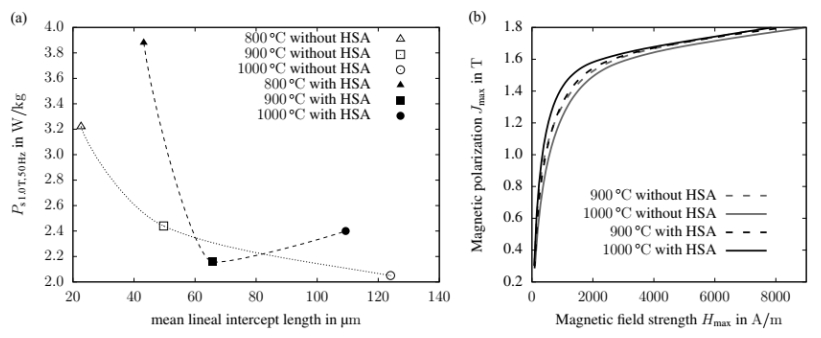
Figures(9) / Tables(1)

Rudolf Kawalla, Anett Stöcker, Ulrich Prahl, Xuefei Wei, Jens Dierdorf, Gerhard Hirt, Martin Heller, Stefan Roggenbuck, Sandra Korte-Kerzel, Hannes Alois Weiss, Philipp Tröber, Lucas Böhm, Wolfram Volk, Nora Leuning, Kay Hameyer. Low-loss FeSi sheet for energy-efficient electrical drives[J]. AIMS Materials Science, 2018, 5(6): 1184-1198. doi: 10.3934/matersci.2018.6.1184







 DownLoad:
DownLoad: