Citation: Florian Thaler , Michael Müller, Robert Spatschek. Oxygen permeation through perovskitic membranes: The influence of steam in the sweep on the permeation performance[J]. AIMS Materials Science, 2016, 3(3): 1126-1137. doi: 10.3934/matersci.2016.3.1126

| [1] |
Czyperek M, Zapp P, Bouwmeester H, et al. (2010) Gas separation membranes for zero-emission fossil power plants: MEM-BRAIN. J Membr Sci 359: 149–159. doi: 10.1016/j.memsci.2010.04.012

|
| [2] | Smart S, Liu S, Serra J, et al. (2014) Perovskite membrane reactors: fundamentals and applications for oxygen production, syngas production and hydrogen processing. In: Gugliuzza A, Basile A, Membranes for Clean and Renewable Power Applications, 1 Eds., Cambridge, UK:Woodhead Publishing, 182–217. |
| [3] |
Benson SJ, Waller D, Kilner JA (1999) Degradation of La0:6Sr0:4Fe0:8O3-δ in carbon dioxide and water atmospheres. J Electrochem Soc 146: 1305–1309. doi: 10.1149/1.1391762
|
| [4] |
Yi J, Feng S, Zuo Y, et al. (2005) Oxygen Permeability and Stability of Sr0:95Co0:8Fe0:2O3-δ in a CO2- and H2O-Containing Atmosphere. Chem Mater 17: 5856–5861. doi: 10.1021/cm051636y
|
| [5] | Yan A, Cheng M, Dong Y, et al. (2006) Investigation of a Ba0.5Sr0.5Co0.8Fe0.2O3 based cathode IT-SOFC: I. The effect of CO2 on the cell performance. Appl Catal B 66: 64–71. |
| [6] |
Arnold M, Wang H, Feldhoff A (2007) Influence of CO2 on the oxygen permeation performance and the microstructure of perovskite-type (Ba0:5Sr0:5)(Co0:8Fe0:2)O3-δ membranes. J Membr Sci 293: 44–52. doi: 10.1016/j.memsci.2007.01.032
|
| [7] |
Waindich A, M?bius A, Müller M (2009) Corrosion of Ba1xSrxCo1yFeyO3-δ and La0:3Ba0:7Co0:2Fe0:2O3-δ materials for oxygen separating membranes under Oxycoal conditions. J Membr Sci 337: 182–187. doi: 10.1016/j.memsci.2009.03.041
|
| [8] |
Engels S, Markus T, Modigell M, et al. (2011) Oxygen permeation and stability investigations on MIEC membrane materials under operating conditions for power plant processes. J Membr Sci 370: 58–69. doi: 10.1016/j.memsci.2010.12.021
|
| [9] |
Song CL, Yi JX (2015) Influence of CO2 on Oxygen Surface Exchange Kinetics of Mixed-Conducting Ba0:5Sr0:5Co0:8Fe0:2O3-δ Oxide. Chinese J Chem Phys 28: 203–205. doi: 10.1063/1674-0068/28/cjcp1412203
|
| [10] |
Wang H, K?lsch P, Schiestel T, et al. (2006) Production of high-purity oxygen by perovskite hollow fiber membranes swept with steam. J Membr Sci 284: 5–8. doi: 10.1016/j.memsci.2006.07.043
|
| [11] |
Leo A, Liu S, Diniz da Costa JC (2011) Production of pure oxygen from BSCF hollow fiber membranes using steam sweep. Sep Purif Technol 78: 220–227. doi: 10.1016/j.seppur.2011.02.006
|
| [12] |
Wang R, Meng B, Meng X, et al. (2015) Highly stable La0:6Sr0:4Co0:2Fe0:2O3-δ hollow fibre membrane for air separation swept by steam or steam mixture. J Membr Sci 479: 232–239. doi: 10.1016/j.memsci.2015.01.006
|
| [13] |
Wang Z, Kathiraser Y, Ang ML, et al. (2015) High Purity Oxygen Production via BBCN Perovskite Hollow Fiber Membrane Swept by Steam. Ind Eng Chem Res 54: 6371–6377. doi: 10.1021/acs.iecr.5b01183
|
| [14] |
Teraoka Y, Zhang H, Furukawa S, et al. (1985) Oxygen permeation through perovskite-type oxides. Chem Lett 14: 1743–1746. doi: 10.1246/cl.1985.1743
|
| [15] |
Baumann S, Serra J, Lobera M, et al. (2011) Ultrahigh oxygen permeation flux through supported Ba0:5Sr0:5Co0:8Fe0:2O3-δ membranes. J Membr Sci 377: 198–205. doi: 10.1016/j.memsci.2011.04.050
|
| [16] |
Mogensen M, Sammes N, Tompsett G (2000) Physical, chemical and electrochemical properties of pure and doped ceria. Solid State Ionics 129: 63–94. doi: 10.1016/S0167-2738(99)00318-5
|
| [17] |
Ten Elshof J, Bouwmeester H, Verweij H (1995) Oxygen transport through La1xSrxFeO3 membranes. I. Permeation in air/He gradients. Solid State Ionics 81: 97–109. doi: 10.1016/0167-2738(95)00177-8
|
| [18] |
Sunarso J, Baumann S, Serra J, et al. (2008) Mixed ionicelectronic conducting (MIEC) ceramicbased membranes for oxygen separation. J Membr Sci 320: 13–41. doi: 10.1016/j.memsci.2008.03.074
|
| [19] |
Shao Z, Yang W, Cong Y, et al. (2000) Investigation of the permeation behavior and stability of a Ba0:5Sr0:5Co0:8Fe0:2O3-δ oxygen membrane. J Membr Sci 172: 177–188. doi: 10.1016/S0376-7388(00)00337-9
|
| [20] |
Serra J, Garcia-Fayos J, Baumann S, et al. (2013) Oxygen permeation through tape-cast asymmetric all-La0:6Sr0:4Co0:2Fe0:2O3-δ membranes. J Membr Sci 447: 297–305. doi: 10.1016/j.memsci.2013.07.030
|
| [21] | Goldschmidt, V (1926) Die Gesetze der Krystallochemie. Die Naturwissenschaften 21: 477–485. |
| [22] |
Bhalla, A, Ruyan G, and Rustum R (2000) The perovskite structure - a review of its role in ceramic science and technology. Mater Res Innovations 4: 3–26. doi: 10.1007/s100190000062
|
| [23] | Bouwmeester H, Burggraaf A (1996) Chapter 10 – Dense ceramic membranes for oxygen separation. In: Burggraaf, A. J. and Cot, L. , Membrane Science and Technology Series, 4, Amsterdam, NL: Elsevier, 435–528. |
| [24] | Huggett L, Piper L (1966) Materials technology in steam reforming processes : proceedings (ed. C. Edeleanu). Materials Technology Symposium Proceedings(1964 : Billingham Eng.) Oxford ; New York : Symposium Publications Division, Pergamon Press, 337. |
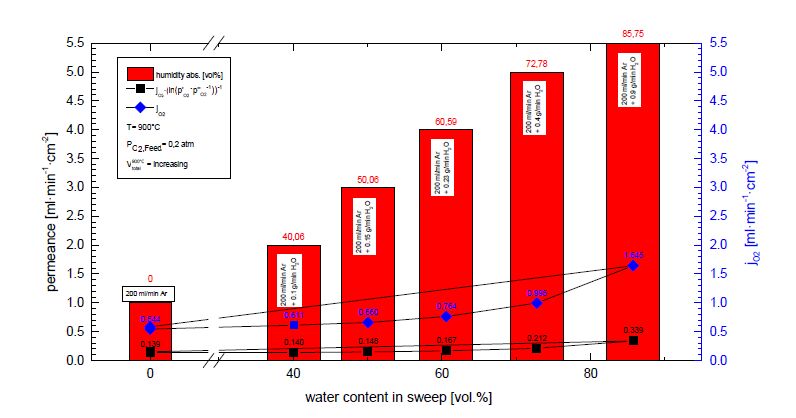
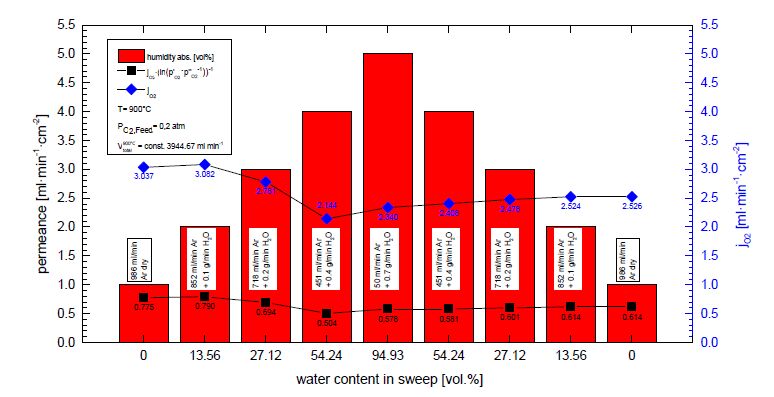

Figures(7)

Florian Thaler , Michael Müller, Robert Spatschek. Oxygen permeation through perovskitic membranes: The influence of steam in the sweep on the permeation performance[J]. AIMS Materials Science, 2016, 3(3): 1126-1137. doi: 10.3934/matersci.2016.3.1126







 DownLoad:
DownLoad: