Citation: Carlos Garcia, Teresa Hernandez, Maria D Coll, Sara Ondoño. Organic amendments for soil restoration in arid and semiarid areas: a review[J]. AIMS Environmental Science, 2017, 4(5): 640-676. doi: 10.3934/environsci.2017.5.640

| [1] | Mol G, Keesstra SD (2012) Soil science in a changing world. Curr Opin Environ Sust 4: 473-477. |
| [2] | Keesstra SD, Bouma J, Wallinga J, et al. (2016) The significance of soils and soil science towards realization of the United Nations Sustainable Development Goals, SOIL 2: 111-128. |
| [3] |
Bastida F, Moreno JL, Hernández T, et al. (2006) Microbiological degradation index of soils in a semiarid climate. Soil Biol Biochem 38: 3463-3473. doi: 10.1016/j.soilbio.2006.06.001

|
| [4] |
Adhikari K, Hartemink AE (2016) Linking soils to ecosystem services-A global review. Geoderma 262: 101-111. doi: 10.1016/j.geoderma.2015.08.009
|
| [5] |
Rasmussen C, Southard RJ, Howarth WR (2006) Mineral control of organic carbon mineralization in a range of temperate conifer forest soils. Global Change Biol 12: 834-847. doi: 10.1111/j.1365-2486.2006.01132.x
|
| [6] |
Lal R, Follett RF, Kimble JM, et al. (2003) Achieving soil carbon sequestration in the United States: a challenge to the policy makers. Soil Sci 168: 827-845. doi: 10.1097/01.ss.0000106407.84926.6b
|
| [7] |
Lal R (2004) Soil carbon sequestration to mitigate climate change. Geoderma 123: 1-22. doi: 10.1016/j.geoderma.2004.01.032
|
| [8] |
Bai ZG, Dent DL, Olsson L, et al. (2008) Proxy global assessment of land degradation. Soil Use Manage 24: 223-234. doi: 10.1111/j.1475-2743.2008.00169.x
|
| [9] | COM 2002, 179. Communication from the Commission to the Council and European Parliament, The European Economic and Social Committee and the Committee of the Regions: Towards a thematic strategy for soil protection. Available from: http:// Europa.eu.int/eur-lex/en/com/pdf/2002/com2002_0170en01.pdf. |
| [10] | Council Directive 2004/35/EC of the European Parliament and of the Council of 21 April 2004 on environmental liability with regard to the prevention and remedying of environmental damage. |
| [11] |
Bastida F, Kandeler E, Moreno JL, et al. (2008) Application of fresh and composted organic wastes modifies structure, size and activity XE "activity" of soil microbial community under semiarid climate. Appl Soil Ecol 40: 318-329. doi: 10.1016/j.apsoil.2008.05.007
|
| [12] |
Lal R (2011) Sequestering carbon in soils of agro-ecosystems. Food Policy 36: 33-39. doi: 10.1016/j.foodpol.2010.12.001
|
| [13] |
Kaiser K, Guggenberger G (2003) Mineral surfaces and soil organic matter. Eur J Soil Sci 54: 219-236. doi: 10.1046/j.1365-2389.2003.00544.x
|
| [14] |
Kalbitz K, Schwesiga D, Rethemeyerb J, et al. (2005) Stabilization of dissolved organic matter by sorption to the mineral soil. Soil Biol Biochem 37: 1319-1331. doi: 10.1016/j.soilbio.2004.11.028
|
| [15] | Six J, Jastrow, JD (2002) Organic matter turnover. Encyclopedia of Soil Science. Marcel Dekker, Inc. |
| [16] |
Pulleman MM, Marinissen JCY (2004) Physical protection of mineralizable C in aggregates from long-term pasture and arable soil. Geoderma 120: 273-282. doi: 10.1016/j.geoderma.2003.09.009
|
| [17] |
García C, Hernández T, Roldán A, et al. (2000) Organic amendment and mycorrhizal inoculation as a practice in afforestation of soils with Pinus halepensis Miller: effect on their microbial activity. Soil Biol Biochem 32: 1173-1181. doi: 10.1016/S0038-0717(00)00033-X
|
| [18] | Tejada M, García C, Gonzalez JL, et al. (2006) Use of organic amendment as a strategy for saline soil remediation: Influence on the physical, Chemical and biological properties of the soil. Soil Biol Biochem 38: 1413-1421. |
| [19] |
Garcia C, Hernandez T (1996) Organic matter in bare soils of the Mediterranean region with semi-arid climate. Arid Soil Res Rehab 10: 31-41. doi: 10.1080/15324989609381418
|
| [20] | Zdruli P, Jones RJA, Montanarella L (2004) Organic matter in the soils of Southern Europe. European Soil Bureau Research Report Nº 15. OOP EC, EUR 21083 EN Luxembourg. |
| [21] | Hernández MT, García E, García C (2015) A strategy for marginal semiarid degraded soil restoration: A sole addition of compost at a high rate. A five-year field experiment. Soil Biol Biochem 89: 61-71. |
| [22] |
Zalidis G, Stamatiadis S, Takavaoglou V, et al. (2002) Impacts of agricultural practices on soil and water quality in the Mediterranean region and proposed assessment methodology. Agric Ecosyt Environ 88: 137-146. doi: 10.1016/S0167-8809(01)00249-3
|
| [23] |
Dunjó G, Pardini G, Gispert M (2003) Land use change effects on abandoned terraced soils in a Mediterranean catchment, NE Spain. Catena 52: 23-37. doi: 10.1016/S0341-8162(02)00148-0
|
| [24] |
Komatsuzaki M, Ohta H (2007) Soil management practices for sustainable agro-ecosystems. Sustain Sci 2: 103-120. doi: 10.1007/s11625-006-0014-5
|
| [25] |
Steenwerth KL, Hodson AK, Bloom AJ, et al. (2014) Climate-smart agriculture global research agenda: scientific basis for action. Agric Food Sec 3:11. doi: 10.1186/2048-7010-3-11
|
| [26] |
Calleja-Cervantes ME, Fernández-González AJ, Irigoyen I, et al. (2015) Thirteen years of continued application of composted organic wastes in a vineyard modify soil quality characteristics. Soil Biol Biochem 90: 241-259. doi: 10.1016/j.soilbio.2015.07.002
|
| [27] |
Diacono M, Montemurro F (2010) Long-term effects of organic amendments on soil fertility. A review. Agron Sustain Dev 30: 401-422. doi: 10.1051/agro/2009040
|
| [28] |
Bastida F, Selevsek N, Torres IF, et al. (2015) Soil restoration with organic amendments: linking cellular functionality and ecosystem processes. Sci Rep 5: 15550. doi: 10.1038/srep15550
|
| [29] |
Nkoa K (2014) Agricultural benefits and environmental risks of soil fertilization with anaerobic digestates: a review. Agron Sustain Dev 34: 473-492. doi: 10.1007/s13593-013-0196-z
|
| [30] |
Antolin MC, Pascaul I, Garcia C, et al. (2005) Growth, yield and solute content of barley in soils treated with sewage sludge under semiarid Mediterranean conditions. Field Crops Res 94: 224-237. doi: 10.1016/j.fcr.2005.01.009
|
| [31] |
Debiase G, Montemurro F, Fioreb A, et al. (2016) Organic amendment and minimum tillage in winter wheat grown in Mediterranean conditions: Effects on yield performance, soil fertility and environmental impact. Eur J Agron 75: 149-157. doi: 10.1016/j.eja.2015.12.009
|
| [32] |
Marron N (2015) Agronomic and environmental effects of land application of residues in short-rotation tree plantations: a literature review. Biomass Bioenerg 81: 378-400. doi: 10.1016/j.biombioe.2015.07.025
|
| [33] | Scotti R, Bonanomi G, Scelza R, et al. (2015) Organic amendments as sustainable tool to recovery fertility in intensive agricultural systems. J Soil Sci Plant Nutr 15: 333-352. |
| [34] |
Scotti R, Pane C, Spaccini R, et al. (2016) On-farm compost: a useful tool to improve soil quality under intensive farming systems. Appl Soil Ecol 107: 13-23. doi: 10.1016/j.apsoil.2016.05.004
|
| [35] | Hernandez MT, Mataix-Solera J, Lichner L, et al. (2007) Assessing the microbiological, biochemical, soil-physical and hydrological effects of amelioration of degraded soils in semiarid Spain. Biologia, 62: 542-546. |
| [36] | Zhang P, Wei T, Jia Z, et al. (2014) Effects of straw incorporation on soil organic matter and soil water-stable aggregates content in semiarid regions of Northwest China, PLoS ONE, 9: e92839. |
| [37] |
Plaza C, Hernández D, García-Gil JC, et al. (2004) Microbial activity in pig slurry-amended soils under semiarid conditions. Soil Biol Biochem 36: 1577-1585. doi: 10.1016/j.soilbio.2004.07.017
|
| [38] |
Tejada M, Hernández T, García C (2009) Soil restoration using composted plant residues: Effects on soil properties. Soil Till Res 102: 109-117. doi: 10.1016/j.still.2008.08.004
|
| [39] |
Ros M, García C, Hernández MT (2007) Pig slurry composts as substitute for mineral fertlizers in horticultural crops. effects on soil characteristics. Renew Agric Food Syst 22: 1-9. doi: 10.1017/S1742170507001500
|
| [40] | Garcia-Ruiz R, Gomez-Munoz B, Hatch DJ, et al. (2012) Soil mineral N retention and N2O emissions following combined application of 15N-labelled fertiliser and weed residues. Rapid Commun Mass Sp 26: 2379-2385. |
| [41] |
Hernández MT, Chocano C, Moreno JL, et al. (2014) Towards a more sustainable fertilization: Combined use of compost and inorganic fertilization for tomato cultivation. Agric Ecosyst Environ 196: 178-184. doi: 10.1016/j.agee.2014.07.006
|
| [42] | Baldi E, Toselli M, Marangoni B (2010) Organic fertilization leads to increased peach root production and lifespan. Tree Physiol 11: 1373-1382. |
| [43] |
Eghball B (2000) Nitrogen mineralization from field-applied beef cattle feedlot manure or compost. Soil Sci Soc Am J 64: 2024-2030. doi: 10.2136/sssaj2000.6462024x
|
| [44] |
Basso B, Ritchie JT (2005) Impact of compost, manure and inorganic fertilizer on nitrate leaching and yield for a 6-year maize–alfalfa rotation in Michigan. Agric Ecosyst Environ 108: 329-341. doi: 10.1016/j.agee.2005.01.011
|
| [45] |
Rathod DD, Rathod PH, Patel KP (2013) Integrated use of organic and inorganic inputs in wheat-fodder maize cropping sequence to improve crop yields and soil properties. Arch Agron Soil Sci 59: 1439-1455. doi: 10.1080/03650340.2012.724171
|
| [46] | Hirzel J, Salazar F (2011) Uso de enmiendas orgánicas como Fuente de fertilización en cultivos. Fertilización de cultivos en Chile. In: Hirzel, J., Ed., Colección libros INIA, Nº 28. Chillán 2011. |
| [47] | Perez-Romero LF, Arroyo F, Santamaria C, et al. (2014) Growth, phenology and fruit set of Prunus armeniaca L. (cv. Ninfa) grafted on two rootstocks in organic and conventional management. Hort Sci 41: 101-106. |
| [48] |
Lal R (2003) Soil erosion and the global carbon budget. Environ Int 29: 437-450. doi: 10.1016/S0160-4120(02)00192-7
|
| [49] | Kizilkaya K, Bayrakli B (2005) Effects of N-enriched sewage sludge on soil enzyme activities. Appl Soil Ecol 30: 192-202. |
| [50] |
Carlson J, Saxena J, Basta N, et al. (2015) Application of organic amendments to restore degraded soil: effects on soil microbial properties. Environ Monit Assess 187: 109. doi: 10.1007/s10661-015-4293-0
|
| [51] | US EPA (2007) The use of soil amendments for remediation, revitalization and reuse. Available form: http://www.epa.gov. |
| [52] |
O'Connor GA, Elliott NT, Basta RK, et al. (2005) Sustainable Land Application. J Environ Qual 34: 7-17. doi: 10.2134/jeq2005.0007
|
| [53] |
Pascual JA, Garcia C, Hernandez T (1999) Lasting microbiological and biochemical effects of the addition of municipal solid waste on an arid soil. Biol Fert Soils 30: 1-6. doi: 10.1007/s003740050579
|
| [54] | Otten L (2001) Wet-dry composting of organic municipal solid waste: current status in Canada. Can J Civil Eng 28: 124-130. |
| [55] |
Wolkowski RP (2003) Nitrogen management considerations for land spreading municipal solid waste compost. J Environ Qual 32: 1844-1850. doi: 10.2134/jeq2003.1844
|
| [56] | Khalil AI, Hassouna MS, El-Ashqar HMA, et al. (2011) Changes in physical, chemical and microbial parameters during the composting of municipal sewage sludge. World J Microb Biot 26: 2117-2122. |
| [57] | Wu D, Liu P, Luo Y, et al. (2010) Nitrogen transformations during co-composting of herbal residues, spent mushrooms, and sludge. J Zhejiang Univ Sci B Biomed Biotechnol 11: 497-505. |
| [58] | Soumaré M, FMG Tack, MG Verloo (2003) Effects of a municipal solid waste compost and mineral fertilization on plant growth in two tropical agricultural soils of Mali. Bioresour Technol 86: 15-20. |
| [59] |
García-Gil JC, Plaza C, Soler-Rovira P, et al. (2000) Long-term effects of municipal solid waste compost application on soil enzyme activities and microbial biomass. Soil Biol Biochem 32: 1907-1913. doi: 10.1016/S0038-0717(00)00165-6
|
| [60] |
Tejada M, Gonzalez JL, García-Martínez AM, et al. (2008) Application of a green manure and green manure composted with beet vinasse on soil restoration: Effects on soil properties. Bioresour Technol 99: 4949-4957. doi: 10.1016/j.biortech.2007.09.026
|
| [61] |
García-Martínez AM, Diaz A, Tejada M, et al. (2010) Enzymatic production of an organic soil biostimulant from wheat-condensed distiller solubles: Effects on soil biochemistry and biodiversity. Process Biochem 45: 1127-1133. doi: 10.1016/j.procbio.2010.04.005
|
| [62] |
Parrado J, Bautista J, Romero EJ, et al. (2008) Production of a carob enzymatic extract: potential use as a biofertilizer. Bioresour Technol 99: 2312-2318. doi: 10.1016/j.biortech.2007.05.029
|
| [63] |
Parrado J, Escudero ML, Friaza V (2007) Enzymatic vegetable extract with bio-active components: influence of fertiliser on the color and anthocyanins of red grapes. J Sci Food Agric 87: 2310-2318. doi: 10.1002/jsfa.2989
|
| [64] |
Tejada M, Benítez C, Gómez I, et al. (2011) Use of biostimulants on soil restoration: Effects on soil biochemical properties and microbial community. App Soil Ecol 49: 11-17. doi: 10.1016/j.apsoil.2011.07.009
|
| [65] |
García C, Hernández T, Costa F (1994) Microbial activity in soils under Mediterranean environmental conditions. Soil Biol Biochem 26: 1185-1191. doi: 10.1016/0038-0717(94)90142-2
|
| [66] |
Prayogo C, Jones JE, Baeyens J, et al. (2014) Impact of biochar on mineralisation of C and N from soil and willow litter and its relationship with microbial community biomass and structure. Biol Fert Soils 50: 695-702. doi: 10.1007/s00374-013-0884-5
|
| [67] | Mankasingh U, Choi PC, Ragnarsdottir V (2009) Biochar application in Tamil Nadu and the global food crisis. In Goldschmitdt Conference-Challenges to our Volatile Planet, in Davos (Switzerland), Jun 21-26. |
| [68] |
Singh R, Babu JN, Kumar R, et al. (2015) Multifaceted application of crop residue biochar as a tool for sustainable agricultura: en ecological perspective. Ecol Eng 77: 324-347. doi: 10.1016/j.ecoleng.2015.01.011
|
| [69] | Uchimiya M, Lima IM, Thomas Klasson K, et al. (2010) Immobilization of heavy metal ions (Cu II, Cd II, Ni II, and Pb II) by broiler litter-derived biochars in water and soil. J Agric Food Chem 58: 5538-5544. |
| [70] |
Kuppusamy S, Thavamani P, Megharaj M, et al. (2016) Agronomic and remedial benefits and risks of applying biochar to soil: Current knowledge and future research directions. Environ Int 87: 1-12. doi: 10.1016/j.envint.2015.10.018
|
| [71] | Novak J, Lima I, Xing B (2009) Characterization of designer biochar produced at different temperatures and their effects on a loamy sand. Annals Environ Sci 3: 195-206. |
| [72] |
Laird D, Fleming P, Wang B, et al. (2010) Biochar impact on nutrient leaching from a Midwestern agricultural soil. Geoderma 158: 436-442. doi: 10.1016/j.geoderma.2010.05.012
|
| [73] |
Chen J, Liu X, Zheng J, et al. (2013) Biochar soil amendment increased bacterial but decreased fungal gene abundance with shifts in community structure in a slightly acid rice paddy from Southwest China. Appl Soil Ecol 71: 33-44. doi: 10.1016/j.apsoil.2013.05.003
|
| [74] |
Jindo K, Sánchez-Monedero MA, Hernández T, et al. (2012) Biochar influences the microbial community structure during manure composting with agricultural wastes. Sci Total Environ 416: 476-481. doi: 10.1016/j.scitotenv.2011.12.009
|
| [75] |
Khodadad CL, Zimmerman AR, Green SJ, et al. (2011) Taxa-specific changes in soil microbial community composition induced by pyrogenic carbon amendments. Soil Biol Biochem 43: 385-392. doi: 10.1016/j.soilbio.2010.11.005
|
| [76] |
Grossman JM, O´Neil BE, Tsai SM, et al. (2010) Amazonian anthrosols support similar microbial communities that differ distinctly from those extant in adjacent, unmodified soils of the same mineralogy. Microb Ecol 60: 192-205. doi: 10.1007/s00248-010-9689-3
|
| [77] |
Nicolás C, Hernández T, García C (2012) Organic amendments as strategy to increase organic matter in particle-size fractions of a semi-arid soil. Appl Soil Ecol 57: 50-58. doi: 10.1016/j.apsoil.2012.02.018
|
| [78] |
Lal R (2003) Soil erosion and the global carbon budget. Environ Int 29: 337-450. doi: 10.1016/S0160-4120(02)00166-6
|
| [79] | Krull ES, Skjemstad JO, Baldock JA (2004) Functions of soil organic matter and the effect on soil properties. In Residue, Soil Organic Carbon and Crop Performance, CSIRO Land and Water, PMB2, Glen Osmond SA 5064. GRDC Project No CSO 00029. |
| [80] |
Zimmerman M, Leifeld J, Smchmidt MWI, et al. (2007) Measured soil organic matter fractions can be related to pools in the Roth C model. Eur J Soil Sci 58: 658-667. doi: 10.1111/j.1365-2389.2006.00855.x
|
| [81] |
García-Franco N, Martínez-Mena M, Goberna, M, et al. (2015) Changes in soil aggregation and microbial community structure control carbon sequestration after afforestation of semiarid shrublands. Soil Biol Biochem 87: 110-121. doi: 10.1016/j.soilbio.2015.04.012
|
| [82] |
García C, Hernández T, Roldán A, et al. (2002) Effect of plant cover decline on chemical microbiological parameters under Mediterranean climate. Soil Biol Biochem 34: 635-642. doi: 10.1016/S0038-0717(01)00225-5
|
| [83] | Cerdà A, González-Pelayo O, Giménez-Morera A, et al. (2016) The use of barley straw residues to avoid high erosion and runoff rates on persimmon plantations in Eastern Spain under low frequency-high magnitude simulated rainfall events. Soil Res 54: 154-165. |
| [84] |
Garcia C, Hernandez T, Costa F (1992) Variation in some chemical parameters and organic matter in soils regenerated by the addition of municipal solid-waste. Environ Manag 16: 763-768. doi: 10.1007/BF02645666
|
| [85] |
Pascual JA, García C, Hernández T, et al. (1997) Changes in the microbial activity of an arid soil amended with urban organic wastes. Biol Fertil Soils 24: 429-434. doi: 10.1007/s003740050268
|
| [86] |
Bastida F, Moreno JL, Garcia C, et al. (2007) Addition of urban waste to semiarid degraded soil: Long-term effect. Pedosphere 17: 557-567. doi: 10.1016/S1002-0160(07)60066-6
|
| [87] |
García-Díaz A, Allas RB, Gristina L, et al. (2016) Carbon input threshold for soil carbon budget optimization in eroding vineyards. Geoderma 271: 144-149. doi: 10.1016/j.geoderma.2016.02.020
|
| [88] | Bastida F, Hernandez T, Albaladejo J, et al. (2013) Phylogenetic and functional changes in the microbial community of long-term restored soils under semiarid climate. Soil Biol Biochem 65, 12-21. |
| [89] | Council Directive 96/61/EC of 24 September 1996 concerning integrated pollution prevention and control (IPPC). |
| [90] |
Agnello AC, Huguenot D, Van Hullebusch ED, et al. (2014) Enhanced phytoremediation: A review of low molecular weight organic acids and surfactants used as amendments. Crit Rev Env Sci Tec 44: 2531-2576. doi: 10.1080/10643389.2013.829764
|
| [91] |
Wilcke W (2007) Global patterns of polycyclic aromatic hydrocarbons (PAHs) in soil. Geoderma 141: 157-166. doi: 10.1016/j.geoderma.2007.07.007
|
| [92] | Boitsov S, Jensen HKB, Klungsøyr J (2009) Natural background and anthropogenic inputs of polycyclic aromatic hydrocarbons (PAH) in sediments of South-Western Barents Sea Mar. Environ Res 68: 236-245. |
| [93] |
Joner EJ, Johansen A, Loibner AP, et al. (2001) Rhizosphere effects on microbial community structure and dissipation and toxicity of polycyclic aromatic hydrocarbons (PAHs) in spiked soil. Environ Sci Technol 35: 2773-2777. doi: 10.1021/es000288s
|
| [94] | Haritash AK, Kaushik CP (2009) Biodegradation aspects of polycyclic aromatic hydrocarbons (PAHs): A review. J Hazard Mater 169: 1-15. |
| [95] | Thapa B, Kumar KCA, Ghimire A (2012) A review on bioremediation of petroleum hydrocarbon contaminants in soil. Kathmandu Univ. J Sci Eng Technol 8: 164-170. |
| [96] |
Kanaly RA, Harayama S (2010) Advances in the field of high-molecular weight polycyclic aromatic hydrocarbon biodegradation by bacteria. Microb Biotechnol 3: 136-164. doi: 10.1111/j.1751-7915.2009.00130.x
|
| [97] | Loick N, Hobbs PJ, Hale MCD, et al. (2012) Bioremediation of poly-aromatic hyrdocarbon (PAH)-contaminated Soil by Composting. Crit Rev Env Sci Tec 39: 271-332. |
| [98] |
García-Delgado C, D'Annibale A, Pesciaroli L, et al. (2015) Implications of polluted soil biostimulation and bioaugmentation with spent mushroom substrate (Agaricus bisporus) on the microbial community and polycyclic aromatic hydrocarbons biodegradation. Sci Tot Environ 508: 20-28. doi: 10.1016/j.scitotenv.2014.11.046
|
| [99] |
Kästner M, Miltner A (2016) Application of compost for effective bioremediation of organic contaminants and pollutants in soil. Appl Microbiol Biot 100: 3433-3449. doi: 10.1007/s00253-016-7378-y
|
| [100] |
Gan S, Lau EV, Ng HK (2009) Remediation of soils contaminated with polycyclic aromatic hydrocarbons (PAHs). J Hazar Mater 172: 532-539. doi: 10.1016/j.jhazmat.2009.07.118
|
| [101] | Samantha L Clay (2014) Identifying the fate of petroleum hydrocarbons released into the environment and their potential biodegradation using stable carbon isotopes and microbial lipid analysis. (Thesis) Master, McMaster University, Hamilton, Ontario. |
| [102] | Ros M, Rodríguez I, García C, et al. (2010) Microbial communities involved in the bioremediation of an aged recalcitrant hydrocarbon polluted soil by using organic amendments. Bioresour Technol 101:6916-6923. |
| [103] | Said-Pullicino D, Gigliotti G, Vella AJ (2004) Environmental fate of triasulfuron in soils amended with municipal waste compost. J Environ Qual 33: 1743-1751. |
| [104] | Purnomo AS, Mori T, KameiI N, et al. (2010) Application of mushroom waste medium from Pleurotus ostreatusfor bioremediation of DDT-contaminated soil. Int Biodeter Biodegr 64: 397-402. |
| [105] |
Jaspers CJ, Ewbank G, McCarthy AJ, et al. (2002) Successive rapid reductive dehalogenation and mineralization of pentachlorophenol by the indigenous microflora of farmyard manure compost. J Appl Microb 92: 127-133. doi: 10.1046/j.1365-2672.2002.01520.x
|
| [106] |
Wang H, Brown SL, Magesan GN, et al. (2008) Technological options for the management of biosolids. Environ Sci Pollut Res 15: 308-317. doi: 10.1007/s11356-008-0012-5
|
| [107] |
Guerrero C, Gomez I, Moral P, et al. (2001) Reclamation of a burned forest soil with municipal waste compost: macronutrient dynamic and improved vegetation cover recovery. Bioresour Technol 76: 221-227. doi: 10.1016/S0960-8524(00)00125-5
|
| [108] |
Juwarkar AA, Jambhulkar HP (2008) Phytoremediation of coal mine spoil dump through integrated biotechnological approach. Bioresour Technol 99: 4732-4741. doi: 10.1016/j.biortech.2007.09.060
|
| [109] |
Kumar GP, Yadav SK, Thawle PR, et al. (2008). Growth of Jatropha curcas onmetal contaminated soil amended with industrial wastes and Azotobacter-a greenhouse study. Bioresour Technol 99: 2078-2082. doi: 10.1016/j.biortech.2007.03.032
|
| [110] | Chiochetta CG, Cotelle S, Masfaraud JF, et al. (2016) Use of agro-industrial organic sludge amendment to remediate degraded soil: chemical and eco(geno)toxicological differences between fresh and stabilized sludge and establishment of application rates. Environ Sci Pollut Res Int 23: 3018-3025. |
| [111] |
Bianchi V, Masciandaro G, Giraldi D, et al. (2008) Enhanced heavy metal phytoextraction from marine dredged sediments comparing conventional chelating agents (Citric Acid and EDTA) with Humic Substances. Water Air Soil Pollut 193: 323-333. doi: 10.1007/s11270-008-9693-0
|
| [112] | EEA environmental statement (2007) EMAS (with data from 2006). Luxembourg: Office for Official Publications of the European Communities. ISSN 1725-9851, ISBN 978-92-9167-936-2. |
| [113] | Jiménez-Cárceles FJ, Egea C, Rodríguez-Caparrós AB, et al. (2006) Contents of nitrogen, ammonium, phosphorus, pesticides and heavy metals, in a salt marsh in the coast of the Mar Menor lagoon (SE Spain). Fresenius Environ Bull 15: 370-378. |
| [114] | Hseu Z, Su SW, Yu HY, et al. (2010) Remediation techniques and heavy metal uptake by different rice varieties in metal‐contaminated soils of Taiwan: New aspects for food safety regulation and sustainable agriculture. Soil Sci Plant Nutr 5: 31-52. |
| [115] | Pardo Martínez-Fernández D, Clemente R, Walker DJ, et al. (2013) The use of olive-mill waste compost to promote the plant vegetation cover in a trace-element-contaminated soil. Environ Sci Pollut Res 21: 1029-1038. |
| [116] | He Z, Shentu J, Yang X, et al. (2015) Heavy metal contamination of soils: sources, indicators, and assessment. J Environ Indic 9:17-18. |
| [117] | Salt DE, Smith RD, Raskin L (1998) Phytoremediation. Ann Rev Plant Phys Plant Mol Biol 49: 643-668. |
| [118] |
Doni S, Macci C, Peruzzi E, et al. (2015) Heavy metal distribution in a sediment phytoremediation system at pilot scale. Ecol Eng 81: 146-157. doi: 10.1016/j.ecoleng.2015.04.049
|
| [119] |
Hueso-González P, Martínez-Murillo JF, Ruiz-Sinoga JDM (2014) The impact of organic amendments on forest soil properties under Mediterranean climatic conditions. Land Degrad Dev 25: 604-612. doi: 10.1002/ldr.2296
|
| [120] | Paz-Ferreiro J, Lu H, Fu S, et al. (2013) Use of phytoremediation and biochar to remediate heavy metal polluted soils: a review. Solid Earth Discuss 5: 2155-2179. |
| [121] |
Luna L, Pastorelli R, Bastida F, et al. (2016) The combination of quarry restoration strategies in semiarid climate induces different responses in biochemical and microbiological soil properties. Appl Soil Ecol 107: 33-47. doi: 10.1016/j.apsoil.2016.05.006
|
| [122] |
Lakhdar A, Rabhi M, Ghnaya T, et al. (2009) Effectiveness of compost use in salt-affected soil. J Hazar Mater 171: 29-37. doi: 10.1016/j.jhazmat.2009.05.132
|
| [123] |
Bhattacharyya R, Kundu S, Prakash V, et al. (2008) Sustainability under combined application of mineral and organic fertilizers in a rainfed soybean-wheat system of the Indian Himalayas. Eur J Agron 28: 33-46. doi: 10.1016/j.eja.2007.04.006
|
| [124] |
Manna MC, Swarup A, Wanjari RH, et al. (2005) Long-term effect of fertilizer and manure application on soil organic carbon storage, soil quality and yield sustainability under sub-humid and semi-arid tropical India. Field Crops Res 93: 264-280. doi: 10.1016/j.fcr.2004.10.006
|
| [125] |
Ros M, Hernández MT, García C (2003) Soil microbial activity after restoration of a semiarid soil by organic amendments. Soil Biol Biochem 35: 463-469. doi: 10.1016/S0038-0717(02)00298-5
|
| [126] |
Garg RN, Pathak H, Das DK, et al. (2005) Use of fly ash and biogas slurry for improving wheat yield and physical properties of the soil. Environ Monit Assess 107: 1-9. doi: 10.1007/s10661-005-2021-x
|
| [127] | Ramulu US (2001) Reuse of municipal sewage and sludge in agriculture. Scientific Publishers, Jodhpur, India. |
| [128] | Parkpain PS, Sirisukhodom AA, Carbonell‐Barrachina (1998) Heavy metals and nutrients chemistry in sewage sludge amended Thai soils. Toxic/Hazar Subst Environ Eng 33: 4573-597. |
| [129] |
Albiach R, Canet R, Pomares F, et al. (2001) Organic matter components and aggregate stability after the application of different amendments to a horticultural soil. Bioresour Technol 76: 125-129. doi: 10.1016/S0960-8524(00)00090-0
|
| [130] | Annabi M, Houot S, Francou C, et al. (2006) Improvement of aggregate stability after urban compost addition in a silty soil, in: Kraft E, Bidlingmaier W, de Bertoldi M, et al. Author, Biological Waste Management, From Local to Global, Proceedings of the International Conference ORBIT2006 /Eckhard Kraft.(Hrsg.) Weimar: Verlag ORBITe.V |
| [131] |
Blanchet G, Gavazov K, Bragazza L, et al. (2016) Responses of soil properties and crop yields to different inorganic and organic amendments in a Swiss conventional farming system. Agric Ecosyst Environ 230: 116-126. doi: 10.1016/j.agee.2016.05.032
|
| [132] |
Jiang MB, Wang XH, Liusui YH, et al. (2017) Variation of soil aggregation and intra-aggregate carbon by long-term fertilization with aggregate formation in a grey desert soil. Catena 149: 437-445. doi: 10.1016/j.catena.2016.10.021
|
| [133] | Peikert B, Schaumann GE, Bibus D, et al. (2017) Effects of olive oil mill wastewater on chemical, microbiological, and physical properties of soil incubated under four different climatic conditions. Biol Fert Soils: 53, 89-102. |
| [134] | Van-Camp L, Bujarrabal B, Gentile AR, et al. (2004) Reports of the Technical Working Groups Established under the Thematic Strategy for Soil Protection, Office for Official Publications of the European Communities, Luxembourg. |
| [135] |
Garcia C, Hernandez T, Albaladejo J (1998) Revegetation in semiarid zones: influence of terracing and organic refuse on microbial activity. Soil Sci Soc Am J 62: 670-76. doi: 10.2136/sssaj1998.03615995006200030018x
|
| [136] | Bastida F, Torres IF, Hernández T, et al. (2013) Can the labile carbon contribute to carbon immobilization in semiarid soils? Priming effects and microbial community dynamics. Soil Biol Biochem 57: 892-902. |
| [137] |
Tejada M, Moreno JL, Hernández MT, et al. (2007) Application of two beet vinasse forms in soil restoration: Effects on soil properties in and environment in southern Spain. Agric Ecosyst Environ 119: 289-298. doi: 10.1016/j.agee.2006.07.019
|
| [138] |
Kiem R, Kögel-Knabner I (2003) Contribution of lignin and polysaccharides to the refractory carbon pool in C-depleted arable soils. Soil Biol Biochem 35: 101-118. doi: 10.1016/S0038-0717(02)00242-0
|
| [139] |
Abiven S, Menasseri S, Chenu C (2009) The effects of organic inputs over time on soil aggregate stability-A literature analysis. Soil Biol Biochem 41: 1-12. doi: 10.1016/j.soilbio.2008.09.015
|
| [140] |
Gentile R, Vanlauwe B, Six J (2011) Litter quality impacts short-but not long-term soil carbon dynamics in soil aggregate fractions. Ecol Appl 21: 695-703. doi: 10.1890/09-2325.1
|
| [141] |
García E, García C, Hernández T (2012) Evaluation of the suitability of using large amounts of urban wastes for degraded arid soil restoration and C fixation. Eur J Soil Sci 63: 650-658. doi: 10.1111/j.1365-2389.2012.01472.x
|
| [142] |
Bhattacharya SS, Ki-Hyun K, Das S, et al. (2016) A review on the role of organic inputs in maintaining the soil carbon pool of the terrestrial ecosystem. J Environ Manage 167: 214-227. doi: 10.1016/j.jenvman.2015.09.042
|
| [143] |
Alburquerque JA, de la Fuente C, Bernal MP (2011) Improvement of soil quality after "alperujo" compost application to two contaminated soils characterised by differing heavy metal solubility. J Environ Manage 92: 733-741. doi: 10.1016/j.jenvman.2010.10.018
|
| [144] | Heidari G, Mohammadi K, Sohrabi Y (2016) Responses of soil microbial biomass and enzyme activities to tillage and fertilization systems in soybean (Glycine max L.) Production. Front Plant Sci 7: 1730. |
| [145] |
Fontaine S, Mariotti A, Abbadie L (2003) The priming effect of organic matter: a question of microbial competition? Soil Biol Biochem 35: 837-843. doi: 10.1016/S0038-0717(03)00123-8
|
| [146] |
Kuzyakov Y, Friedel JK, Stahr K (2000) Review of mechanism and quantification of priming effects. Soil Biol Biochem 32: 1485-1498. doi: 10.1016/S0038-0717(00)00084-5
|
| [147] |
Blagodatskaya E, Kuzyakov Y (2008) Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure: critical review. Biol Fert Soils 45: 115-131. doi: 10.1007/s00374-008-0334-y
|
| [148] |
Saison C, Degrange V, Oliver R, et al. (2006) Alteration and resilience of the soil microbial community following compost amendment: effects of compost level and compost-borne microbial community. Environ Microb 8: 247-257. doi: 10.1111/j.1462-2920.2005.00892.x
|
| [149] |
Ondoño S, Bastida F, Moreno JL (2014) Microbiological and biochemical properties of artificial substrates: A preliminary study of its application as Technosols or as a basis in Green Roof Systems. Ecol Eng 70: 189-199. doi: 10.1016/j.ecoleng.2014.05.003
|
| [150] |
Kandeler E, Tscherko D, Bruce KD, et al. (2000) Structure and function of the soil microbial community in microhabitats of a heavy metal polluted soil. Biol Fertil Soils 32: 390-400. doi: 10.1007/s003740000268
|
| [151] |
Pascual JA, García C, Hernández T, et al, (2000) Soil microbial activity as a biomarker of degradation and remediation processes. Soil Biol Biochem 32: 1877-1883. doi: 10.1016/S0038-0717(00)00161-9
|
| [152] | Alguacil MM, Díaz-Pereira E, Caravaca F, et al. (2009) Increased diversity of arbuscular mycorrhizal fungi in a long-term field experiment via application of organic amendments to a semiarid degraded soil. Appl Environ Microb 75: 4254-4263. |
| [153] |
Fierer N, Jackson RB (2006) The diversity and biogeography of soil bacterial communities. P Nat Acad Sci USA 103: 626-631. doi: 10.1073/pnas.0507535103
|
| [154] | Acosta-Martínez V, Dowd S, Sun Y, et al. (2008) Tag-encoded pyrosequencing analysis of bacterial diversity in a single soil type as affected by management and land use. Soil Biol Biochem 40: 2762-2770. |
| [155] |
Oren A, Steinberger Y (2008) Catabolic profiles of soil fungal communities along a geographic climatic gradient in Israel. Soil Biol Biochem 40: 2578-2587. doi: 10.1016/j.soilbio.2008.05.024
|
| [156] |
Goberna M, Navarro-Cano JA, Valiente-Nauet A, et al. (2014) Abiotic stress tolerance and competition-related traits underlie phylogenetic clustering in soil bacterial communities. Ecol Lett 17: 1191-1201. doi: 10.1111/ele.12341
|
| [157] |
Chaudhry V, Rehman A, Mishra A, et al. (2012) Changes in bacterial community structure of agricultural land due to long-term organic and chemical amendments. Microb Ecol 64: 450-460. doi: 10.1007/s00248-012-0025-y
|
| [158] |
Ben-David EA, Zaady E, Sher Y, et al. (2011) Assessment of the spatial distribution of soil microbial communities in patchy arid and semi-arid landscapes of the Negev Desert using combined PLFA and DGGE analyses. FEMS Microbiol Ecol 76: 492-503. doi: 10.1111/j.1574-6941.2011.01075.x
|
| [159] | Kuske CR, Ticknor LO, Miller ME, et al. (2002) Comparison of soil bacteria communities in rizhospheres of three plant species and the interspaces in an arid grassland. Appl Environ Microbiol 68: 1854-1863. |
| [160] |
Hortal S, Bastida F, Armas C, et al. (2013) Soil microbial community under a nurse-plant species changes in composition, biomass and activity as the nurse grows. Soil Biol Biochem 64: 139-146. doi: 10.1016/j.soilbio.2013.04.018
|
| [161] | Angel R, Soares MIM, Ungar E, et al. (2010) Biogeography of soil archaea and bacteria along a steep precipitation gradient. The ISME Journal 4: 553-563. |
| [162] |
Neilson JW, Quade J, Ortiz M, et al. (2012) Life at the hyperarid margin: novel bacterial diversity in arid soils of the Atacama Desert, Chile. Extremophiles 16: 553. doi: 10.1007/s00792-012-0454-z
|
| [163] | Connon SA, Lester ED, Shafaat HS, et al. (2007). Bacterial diversity in hyperarid Atacama desert soils. J Geophys Res: Biogeosciences 112. |
| [164] |
Drees KP, Nielson JW, Betancourt JL, et al. (2006) Bacterial community structure in the hyperarid core of the Atacama Desert, Chile. Appl Environ Microb 72: 7902-7908. doi: 10.1128/AEM.01305-06
|
| [165] |
Fontaine S, Bardoux G, Abbadie L, et al. (2004) Carbon input to soil may decrease soil carbon content. Ecol Lett 7: 314-320. doi: 10.1111/j.1461-0248.2004.00579.x
|
| [166] | Senesi N, Plaza C (2007) Role of humification processes in recycling organic wastes of various nature and sources as organic amendments. Clean 35: 26-41. |
| [167] | Schnitzer M, Monreal CM (2011) Quo vadis soil organic matter research? A biological link to the chemistry of humification. In: Sparks DL Author, Advances in Agronomy, Burlington: Academic Press, 139-213. |
| [168] | Singh J, Kalamdhad AS (2012) Reduction of heavy metals during composting-A review. Int J Environ Protect 2: 36-43. |
| [169] | Plaza C, Senesi N (2009) The effect of organic matter amendment on native soil humic substances. In: Senesi PM, Huang XB Author, Biophysico-Chemical Processes Involving Natural Nonliving Organic Matter in Environmental System. Hoboken: Wiley & Sons, 147-181. |
| [170] |
Bernal MP, Alburquerque JM, Moral R (2009) Composting of animal manures and chemical criteria for compost maturity assessment-a review. Bioresour Technol 100: 5444-5453. doi: 10.1016/j.biortech.2008.11.027
|
| [171] |
Larney FJ, Hao X (2007) A review of composting as a management alternative for beef cattle feedlot manure in southern Alberta, Canada. Bioresour Technol 98: 3221-3227. doi: 10.1016/j.biortech.2006.07.005
|
| [172] |
Medina J, Monreal C, Barea JM, et al. (2015) Crop residue stabilization and application to agricultural and degraded soils: A review. Waste Manage 42: 41-54. doi: 10.1016/j.wasman.2015.04.002
|
| [173] |
Lynch DH, Voroney RP, Warman PR (2006) Use of 13C and 15N natural abundance techniques to characterize carbon and nitrogen dynamics in composting and in compost-amended soils. Soil Biol Biochem 38: 103-114. doi: 10.1016/j.soilbio.2005.04.022
|
| [174] |
Walter I, Martinez F, Cuevas G (2006) Plant and soil responses to the application of composted MSW in a degraded, semiarid shrubland in central Spain. Compost Sci Util 14: 147-154. doi: 10.1080/1065657X.2006.10702276
|
| [175] | Tittarelli F, Petruzzelli G, Pezzarossa B, et al. (2007) Quality and agronomic use of compost, in: Diaz LF, de Bertoldi M, Bidlingmaier W, et al. Author, Compost science and technology, Waste management, Elsevier Ltd., 119-145. |
| [176] |
Zhang M, Heaney D, Henriquez B, et al. (2006) A four year study on influence of biosolids/MSW compost application in less productive soils in Alberta: nutrient dynamics. Compost Sci Util 14: 68-80. doi: 10.1080/1065657X.2006.10702265
|
| [177] | Fernández-Getino AP, Pérez J, Albarrán MM, et al. (2012) Restoration of abandoned, degraded agricultural soil using composted biosolid: Influence on selected soil properties. Arid Land Res Manage 26: 200-210. |
| [178] |
Fabrizio A, Tambone F, Genevini P (2009) Effect of compost application rate on carbon degradation and retention in soils. Waste Manage 29: 174-179. doi: 10.1016/j.wasman.2008.02.010
|
| [179] |
Ryals R, Silver WL (2013) Effects of organic matter amendments on net primary productivity and greenhouse gas emissions in annual grasslands. Ecol Appl 23: 46-69. doi: 10.1890/12-0620.1
|
| [180] |
Kätterer T, Börjesson G, Kirchmann H (2014) Changes in organic carbon in topsoil and subsoil and microbial community composition caused by repeated additions of organic amendments and N fertilisation in a long-term field experiment in Sweden. Agric Ecosyst Environ 189:110-118. doi: 10.1016/j.agee.2014.03.025
|
| [181] |
Macías F, Camps-Arbestain M (2010) Soil carbon sequestration in a changing global environment. Mitig Adapt Strat Gl 15: 511-529. doi: 10.1007/s11027-010-9231-4
|
| [182] |
Rovira P, Vallejo VR (2002) Mineralization of carbon and nitrogen from plant debris, as affected by debris size and depth of burial. Soil Biol Biochem 34: 327-339. doi: 10.1016/S0038-0717(01)00186-9
|
| [183] |
Don A, Roedenbeck C, Gleixner G (2013) Unexpected control of soil carbon turnover by soil carbon concentration. Environ Chem Letters 11: 407-413. doi: 10.1007/s10311-013-0433-3
|
| [184] |
Odlare M, Pell M, Svensson K (2008) Changes in soil chemical and microbiological properties during 4 years of application of various organic residues. Waste Manage 28: 1246-1253. doi: 10.1016/j.wasman.2007.06.005
|
| [185] |
Wienhold BJ, Andrews SS, Karlen DL (2004) Soil quality: a review of the science and experiences in the USA. Environ Geochem Health 26: 89-95. doi: 10.1023/B:EGAH.0000039571.59640.3c
|
| [186] |
Ge Y, Changrong C, Zhihong X, et al. (2010) Carbon/nitrogen ratio as a major factor for predicting the effects of organic wastes on soil bacterial communities assessed by DNA-based molecular techniques. Environ Sci Pollut R 17: 807-815. doi: 10.1007/s11356-009-0185-6
|
| [187] |
Alburquerque JA, de la Fuente C, Ferre-Costa A, et al. (2012) Assessment of the fertiliser potential of digestates from farm and agro-industrial residues. Biomass Bioenergy 40: 181-189. doi: 10.1016/j.biombioe.2012.02.018
|
| [188] |
Watcharasukarn M, Kaparaju P, Steyer JP, et al. (2009) Screening Escherichia coli, Enteroccocus faecalis, and Clostridium perfringens as indicator organisms in evaluating pathogen-reducing capacity in biogas plants. Microb Ecol 58: 221-230. doi: 10.1007/s00248-009-9497-9
|
| [189] | Boydston RA, Collins HP, Vaughn SF (2008) Response of weeds and ornamental plants to potting soil amended with dried distillers grains. Hort Sci 43: 191-195. |
| [190] | Kulling D, Stadelmann F, Herter U (2001) Sewage Sludge-Fertilizer or Waste? UKWIR Conference, Brussels. |
| [191] |
Lopez-Mosquera ME, Moiron C, Carral E (2000) Use of dairy industry sludge as fertilizer for grassland in northwest Spain: heavy metal levels in the soil and plants. Resour Conserv Recy 30: 95-109. doi: 10.1016/S0921-3449(00)00058-6
|
| [192] | Spencer JL, Guan J (2004) Public health implications related to spread of pathogens in manure from livestock and poultry operations. In: Spencer JFT, Ragout de Spencer AL Author, Public health microbiology: methods and protocols. Humana, Totowa, 503-515. |
| [193] | Hutchison ML, Walters LD, Avery SM, et al. (2005) Analysis of livestock production, waste storage, and pathogen levels and prevalences in farm manures. Appl Environ Microb 71: 1231-1236. |
| [194] |
Bicudo JR, Goyal SM (2003) Pathogens and manure management systems: a review. Environ Technol 24: 115-130. doi: 10.1080/09593330309385542
|
| [195] |
Masse D, Gilbert Y, Topp E (2011) Pathogen removal in farm-scale psychrophilic anaerobic digesters processing swine manure. Bioresour Technol 102: 641-646. doi: 10.1016/j.biortech.2010.08.020
|
| [196] | Angelidaki I, Ellegaard L, Ahring BK (2003) Applications of the anaerobic digestion process. In: Ahring BK Author, Biomethanation II. Berlin: Springer, 1-33. |
| [197] | Slana I, Pribylova R, Kralova A, et al. (2011) Persistence of Mycobacterium avium subsp. Paratubercolosis at a farm-scale biogas plant supplied with manure from paratuberculosis-affected dairy cattle. Appl Environ Microb 77: 3115-3119. |
| [198] |
Schnurer A, Schnurer J (2006) Fungal survival during anaerobic digestion of organic household waste. Waste Manage 26: 1205-1211. doi: 10.1016/j.wasman.2005.09.007
|
| [199] | Council Directive 86/278/EEC of 12 June 1986 on the protection of the environment, and in particular of the soil, when sewage sludge is used in agriculture. |
| [200] |
Eldridge SM, Chan KY, Barchia I, et al. (2009) A comparison of surface applied granulated biosolids and poultry litter in terms of risk to runoff water quality on turf farms in Western Sydney, Australia. Agric Ecosyst Environ 134: 243-250. doi: 10.1016/j.agee.2009.07.007
|
| [201] | Navarro-Pedreño J, Almendro-Candel MB, Jordán-Vidal MM, et al. (2004) Risk areas in the application of sewage sludge on degraded soils in Alicante province (Spain). In: Martin JF, Brebbia CA, Godfrey AE, et al. Author, Geo-Environment. Southampton: WIT Press, 293-302. |
| [202] | Lag-Brotons AJ, Soriano-Disla JM, Gómez I, et al. (2013) Saline irrigation effects on Cynara cardunculus L. plants grown in Mediterranean soils. Hort Sci 48: 762-767. |
| [203] |
Cai H, Chen T, Liu H, et al. (2010) The effect of salinity and porosity of sewage sludge compost on the growth of vegetable seedlings. Sci Hortic 124: 381-386. doi: 10.1016/j.scienta.2010.01.009
|
| [204] |
Pérez-Gimeno A, Navarro-Pedreño J, Almendro-Candel MB, et al. (2016) Environmental consequences of the use of sewage sludge compost and limestone outcrop residue for soil restoration: salinity and trace elements pollution. J Soil Sediment 16: 1012-1021. doi: 10.1007/s11368-015-1288-y
|
| [205] |
Young I, Crawford J (2004) Interactions and self-organisation in the soil-microbe complex. Science 304: 1634-1637. doi: 10.1126/science.1097394
|
| [206] |
Dominati E, Mackay A, Green S, et al. (2014) A soil change-based methodology for the quantification and valuation of ecosystem services from agro-ecosystems: a case study of pastoral agriculture in New Zealand. Ecol Econ 100: 119-129. doi: 10.1016/j.ecolecon.2014.02.008
|
| [207] | Hewitt A, Dominati E, Webb T, et al. (2015) Soil natural capital quantification by the stock adequacy method. Geoderma 241-242: 107-114. |
| [208] |
Mc Bratney A, Field DJ, Koch A (2014) The dimensions of soil security. Geoderma 213: 203-213. doi: 10.1016/j.geoderma.2013.08.013
|
| [209] | Robinson DA, Hockley N, Dominati EJ, et al. (2012) Natural capital, ecosystem services and soil change: why soil science must embrace an ecosystems approach. Vadose Zone J 11: 1-21. |
| [210] | Bouma J (2015) Reaching out from the soil-box in pursuit of soil security. Soil Sci Plant Nutr 61: 556-565. |
| [211] |
Godfray HCJ, Beddington JR, Crute IR, et al. (2010) Food security: the challenge of feeding 9 billion people. Science 327: 812-818. doi: 10.1126/science.1185383
|
| [212] | Montgomery, HL (2010) How is soil made? Crabtree Publishing, New York. |
| [213] |
Zhou P, Sheng H, Lia Y, et al. (2016) Lower C sequestration and N use efficiency by straw incorporation than manure amendment on paddy soils. Agric Ecosyst Environ 219: 93-100. doi: 10.1016/j.agee.2015.12.012
|
| [214] | Smith P, Fang C, Dawson JJC, et al. (2008) Impact of global warming on soil organic carbon. Adv Agron:97: 1-43. |
| [215] |
Houghton RA (2007) Balancing the global carbon budget. Annu Rev Earth Pl Sc. Book Series. 35: 313-347. doi: 10.1146/annurev.earth.35.031306.140057
|
| [216] |
Six J, Frey SD, Thiet RK, et al. (2006) Bacterial and fungal contributions to carbon sequestration in agroecosystems. Soil Sci Soc Am J 70: 555-569. doi: 10.2136/sssaj2004.0347
|
| [217] |
Powlson D, Whitmore A, Goulding K (2011) Soil carbon sequestration to mitigate climate change: a critical re-examination to identify the true and the false. Eur J Soil Sci 62: 42-55. doi: 10.1111/j.1365-2389.2010.01342.x
|
| [218] | Trasar-Ccepeda C, Leirós MC, Gil-Sotres F (2008) Hydrolytic enzyme activities in agricultural and forest soils. Some implications for their use as indicators of soil quality. Soil Biol Biochem 40: 2146-2155. |
| [219] | Shaw LJ, Burns RG (2006) Enzyme activity profiles and soil quality. In: Bloem J, Hopkins DW, Benedetti A Author, Microbiological Methods for Assessing Soil Quality. CABI, Oxfordshire, IK, 158-182. |
| [220] |
Davidson EA, Janssens IA (2006) Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 440: 165-173. doi: 10.1038/nature04514
|
| [221] |
Lehmann J, Rillig MC, Thies J, et al. (2011) Biochar effects on soil biota-A review. Soil Biol Biochem 43: 1812-1836. doi: 10.1016/j.soilbio.2011.04.022
|
| [222] |
Montiel-Rozas MM, Panettieri M, Madejón, P, et al. (2016) Carbon sequestration in restored soils by applying organic amendments. Land Degrad Dev 27: 620-629. doi: 10.1002/ldr.2466
|
| [223] | Yazdanpanah N, Mahmoodabadi M, Cerdá A (2016) The impact of organic amendments on soil hydrology, structure and microbial respiration in semiarid lands. Geoderma 266: 58-65. |
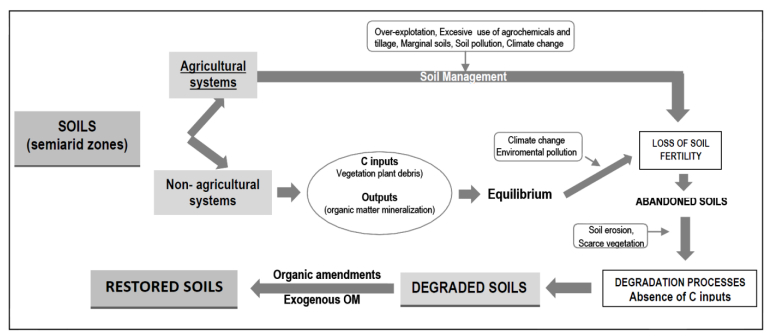
Figures(1) / Tables(3)

Carlos Garcia, Teresa Hernandez, Maria D Coll, Sara Ondoño. Organic amendments for soil restoration in arid and semiarid areas: a review[J]. AIMS Environmental Science, 2017, 4(5): 640-676. doi: 10.3934/environsci.2017.5.640







 DownLoad:
DownLoad: