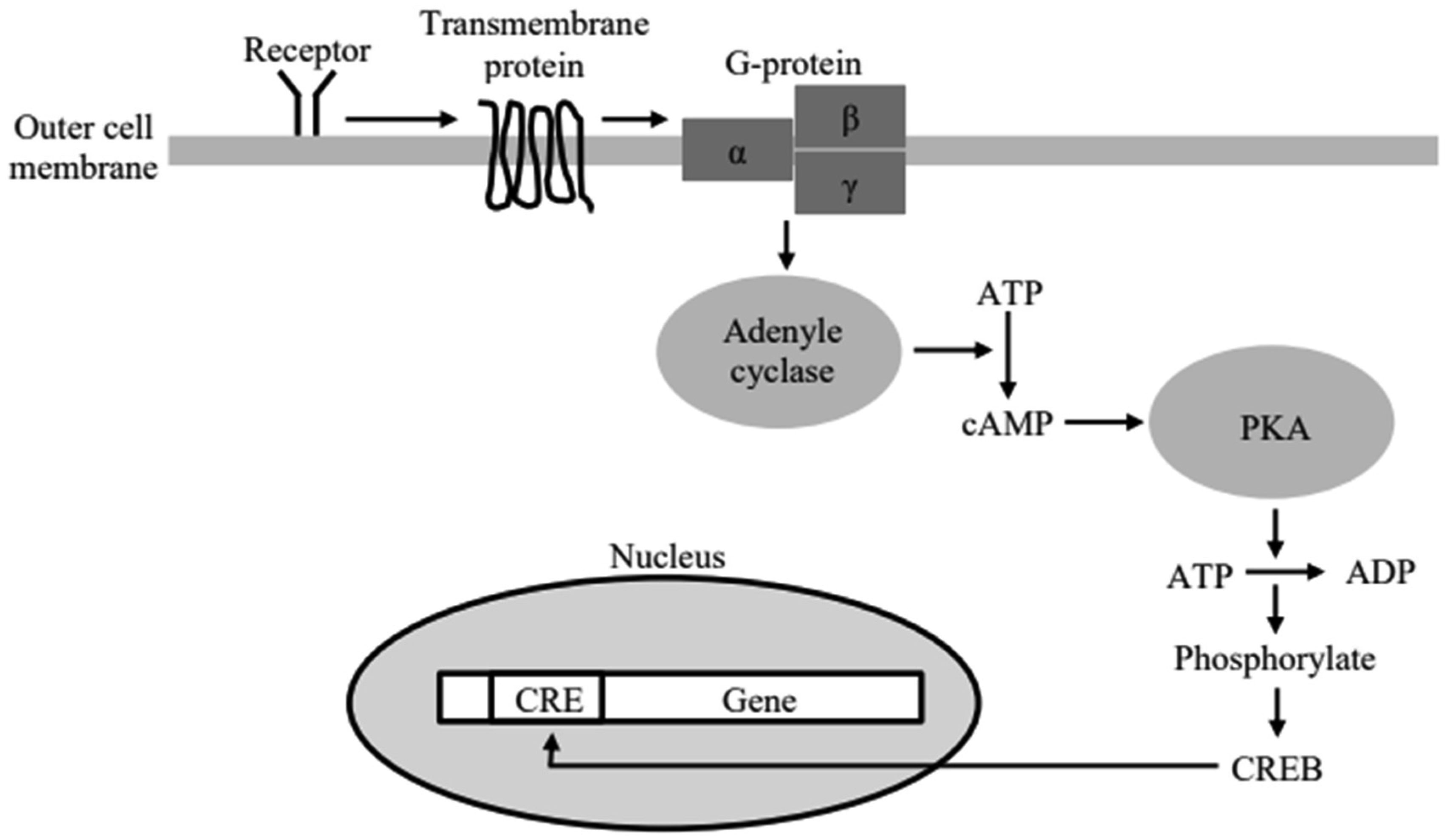
Among the most sought after breakthroughs nowadays to combat computational saturation in the electronic hardware realm, neuromorphic and cytomorphic mimetics of biological structures seem potentially promising. Biological circuits are distinguishable due to their minuscule dimensions and immensely low power consumption; yet they achieve extremely complex and magnificent tasks of life, such as, thinking, memorizing, decision making and self-regulating in response to the surroundings. Low power analog circuit solutions are edged over digital ones as they are inherently noisy and fuzzy like bio-systems. In this paper, an analog circuit equivalent for a well-known biological pathway, cyclic adenosine monophosphate (cAMP), has been proposed, exploiting the fabrication characteristics of an analog transistor. The work demonstrates an application of previously published research of the authors, where it was shown that a single transistor operating in analog mode can mimic some fundamental biological circuit processes like receptor-ligand binding, Michaelis Menten and Hill process reactions. Since biological pathways are chain connections of such reactions, same modular approach can be used to build electronic pathways using those basic transistor circuits. Although the idea of creating silicon life seems far-fetched at this stage, this work supplements the idea of cytomorphic chips which is already gaining interest of bio-engineering community.
Citation: Maria Waqas, Urooj Ainuddin, Umar Iftikhar. An analog electronic circuit model for cAMP-dependent pathway—towards creation of Silicon life[J]. AIMS Bioengineering, 2022, 9(2): 145-162. doi: 10.3934/bioeng.2022011
Among the most sought after breakthroughs nowadays to combat computational saturation in the electronic hardware realm, neuromorphic and cytomorphic mimetics of biological structures seem potentially promising. Biological circuits are distinguishable due to their minuscule dimensions and immensely low power consumption; yet they achieve extremely complex and magnificent tasks of life, such as, thinking, memorizing, decision making and self-regulating in response to the surroundings. Low power analog circuit solutions are edged over digital ones as they are inherently noisy and fuzzy like bio-systems. In this paper, an analog circuit equivalent for a well-known biological pathway, cyclic adenosine monophosphate (cAMP), has been proposed, exploiting the fabrication characteristics of an analog transistor. The work demonstrates an application of previously published research of the authors, where it was shown that a single transistor operating in analog mode can mimic some fundamental biological circuit processes like receptor-ligand binding, Michaelis Menten and Hill process reactions. Since biological pathways are chain connections of such reactions, same modular approach can be used to build electronic pathways using those basic transistor circuits. Although the idea of creating silicon life seems far-fetched at this stage, this work supplements the idea of cytomorphic chips which is already gaining interest of bio-engineering community.

| [1] | Hasan SMR (2008) A novel mixed-signal integrated circuit model for DNA-protein regulatory genetic circuits and genetic state machines. IEEE T Circuits-I 55: 1185-1196. https://doi.org/10.1109/TCSI.2008.925632 |
| [2] | Alam S, Hasan SMR (2013) Integrated circuit modeling of biocellular post-transcription gene mechanisms regulated by microRNA and proteasome. IEEE T Circuits-I 60: 2298-2310. https://doi.org/10.1109/TCSI.2013.2245451 |
| [3] |
Rezaul Hasan SM (2010) A micro-sequenced CMOS model for cell signalling pathway using G-protein and phosphorylation cascade. Int J Comput Appl T 39: 40-45. 
|
| [4] |
Hasan SMR (2010) A digital cmos sequential circuit model for bio-cellular adaptive immune response pathway using phagolysosomic digestion: a digital phagocytosis engine. J Biomed Sci Eng 3: 470-475. https://doi.org/10.4236/jbise.2010.35065
|
| [5] | Ainuddin U, Khurram M (2016) From cell to silicon: Translation of a genetic circuit to finite state machine implementation. IEEE 2016: 376-380. https://doi.org/10.1109/INTECH.2016.7845078 |
| [6] |
Ainuddin U, Khurram M, Hasan SMR (2019) Cloning the λSwitch: digital and markov representations. IEEE T NanoBiosci 18: 428-436. https://doi.org/10.1109/TNB.2019.2908669
|
| [7] | Mandal S, Sarpeshkar R (2009) Log-domain circuit models of chemical reactions. IEEE 2009: 2697-2700. https://doi.org/10.1109/ISCAS.2009.5118358 |
| [8] | Mandal S, Sarpeshkar R (2009) Circuit models of stochastic genetic networks. IEEE 2009: 109-112. https://doi.org/10.1109/BIOCAS.2009.5372073 |
| [9] | Daniel R, Woo SS, Turicchia L, et al. (2011) Analog transistor models of bacterial genetic circuits. IEEE 2011: 333-336. https://doi.org/10.1109/BioCAS.2011.6107795 |
| [10] | Teo JJY, Woo SS, Sarpeshkar R (2015) Synthetic biology: a unifying view and review using analog circuits. IEEE T Biomed Circ S 9: 453-474. https://doi.org/10.1109/TBCAS.2015.2461446 |
| [11] |
Woo SS, Kim J, Sarpeshkar R (2015) A cytomorphic chip for quantitative modeling of fundamental bio-molecular circuits. IEEE T Biomed Circ S 9: 527-542. https://doi.org/10.1109/TBCAS.2015.2446431
|
| [12] |
Achour S, Sarpeshkar R, Rinard MC (2016) Configuration synthesis for programmable analog devices with Arco. ACM SIGPLAN Notices 51: 177-193. https://doi.org/10.1145/2980983.2908116
|
| [13] |
Woo SS, Kim J, Sarpeshkar R (2018) A digitally programmable cytomorphic chip for simulation of arbitrary biochemical reaction networks. IEEE T Biomed Circ S 12: 360-378. https://doi.org/10.1109/TBCAS.2017.2781253
|
| [14] |
Medley JK, Teo J, Woo SS, et al. (2020) A compiler for biological networks on silicon chips. PLoS Comput Biol 16: e1008063. https://doi.org/10.1371/journal.pcbi.1008063
|
| [15] |
Teo JJY, Weiss R, Sarpeshkar R (2019) An artificial tissue homeostasis circuit designed via analog circuit techniques. IEEE T Biomed Circ S 13: 540-553. https://doi.org/10.1109/TBCAS.2019.2907074
|
| [16] | Teo JJY, Kim J, Woo SS, et al. (2019) Bio-molecular circuit design with electronic circuit software and cytomorphic chips. IEEE 2019: 1-4. https://doi.org/10.1109/BIOCAS.2019.8918684 |
| [17] |
Teo JJY, Sarpeshkar R (2020) The merging of biological and electronic circuits. Iscience 23: 101688. https://doi.org/10.1016/j.isci.2020.101688
|
| [18] |
Zeng J, Banerjee A, Kim J, et al. (2019) A novel bioelectronic reporter system in living cells tested with a synthetic biological comparator. Sci Rep 9: 7275. https://doi.org/10.1038/s41598-019-43771-w
|
| [19] |
Zeng J, Teo J, Banerjee A, et al. (2018) A synthetic microbial operational amplifier. ACS Synth Biol 7: 2007-2013. https://doi.org/10.1021/acssynbio.8b00138
|
| [20] |
Kim J, Woo SS, Sarpeshkar R (2018) Fast and precise emulation of stochastic biochemical reaction networks with amplified thermal noise in silicon chips. IEEE T Biomed Circ S 12: 379-389. https://doi.org/10.1109/TBCAS.2017.2786306
|
| [21] |
Banerjee A, Weaver I, Thorsen T, et al. (2017) Bioelectronic measurement and feedback control of molecules in living cells. Sci Rep 7: 12511. https://doi.org/10.1038/s41598-017-12655-2
|
| [22] |
Ahmad W, Rohim RAA, Norhayati Y, et al. (2018) Developing a new dimension of an applied exponential model: application in biological sciences. Eng Technol Appl Sci Res 8: 3130-3134. https://doi.org/10.48084/etasr.2124
|
| [23] |
Ahmad W, Aleng NA, Ali Z, et al. (2018) Statistical modeling via bootstrapping and weighted techniques based on variances. Eng Technol Appl Sci Res 8: 3135-3140. https://doi.org/10.48084/etasr.2126
|
| [24] |
Ahmad W, Rohim RAA, Ismail NH (2019) Estimate outcome value of doubling cell growth using fuzzy regression method. Eng Technol Appl Sci Res 9: 3692-3695. https://doi.org/10.48084/etasr.2467
|
| [25] |
Waqas M, Khurram M, Hasan SM (2017) Bio-cellular processes modeling on silicon substrate: receptor–ligand binding and Michaelis Menten reaction. Analog Integr Circ S 93: 329-340. https://doi.org/10.1007/s10470-017-1044-x
|
| [26] | Waqas M (2019) Integrated circuit models of bio-cellular networks [PhD thesis]. NED University of Engineering & Technology, Karachi . http://173.208.131.244:9060/xmlui/handle/123456789/5232 |
| [27] |
Waqas M, Khurram M, Hasan SMR (2020) Analog electronic circuits to model cooperativity in hill process. Mehran Univ Res J Eng Technol 39: 678-685. https://doi.org/10.22581/muet1982.2004.01
|
| [28] | Zhu Y, Li Y, Zeng N, et al. (2012) Design and analysis of genetic regulatory networks with electronic circuit ideas. IEEE 2012: 2046-2049. https://doi.org/10.1109/ICICEE.2012.544 |
| [29] | Berridge MJ (2007) Cell Signalling Biology: Module 2 Cell Signalling Pathways. Portland Press. |
| [30] | Berridge MJ (2007) Cell Signalling Biology: Module 1 Introduction. Portland Press. |
| [31] | Alberts B, Johnson A, Lewis J, et al. (2002) Molecular Biology of the Cell. New York: Garland Science, Taylor and Francis Group. |
| [32] | Lehninger AL (2004) Lehninger Principles of Biochemistry: David L. Nelson, Michael M. Cox. New York: Recording for the Blind & Dyslexic. |
| [33] | Mayadevi M, Archana GM, Prabhu RR, et al. (2012) Molecular mechanisms in synaptic plasticity. Neuroscience-Dealing With Frontiers . Croatia: IntechOpen 295-330. https://doi.org/10.5772/36928 |
| [34] |
Alberini CM (2009) Transcription factors in long-term memory and synaptic plasticity. Physiol Rev 89: 121-145. https://doi.org/10.1152/physrev.00017.2008
|
| [35] |
Williamson T, Schwartz JM, Kell DB, et al. (2009) Deterministic mathematical models of the cAMP pathway in Saccharomyces cerevisiae. BMC Syst Biol 3: 70. https://doi.org/10.1186/1752-0509-3-70
|
| [36] |
Boyle J (2005) Lehninger principles of biochemistry: Nelson, D., and Cox, M. Biochem Mol Biol Educ 33: 74-75. https://doi.org/10.1002/bmb.2005.494033010419
|
| [37] | Berg JM, Tymoczko JL, Stryer L (2002) The Michaelis-Menten model accounts for the kinetic properties of many enzymes. Biochemistry 5: 319-330. |




Figures(5)

Maria Waqas, Urooj Ainuddin, Umar Iftikhar. An analog electronic circuit model for cAMP-dependent pathway—towards creation of Silicon life[J]. AIMS Bioengineering, 2022, 9(2): 145-162. doi: 10.3934/bioeng.2022011







 DownLoad:
DownLoad: