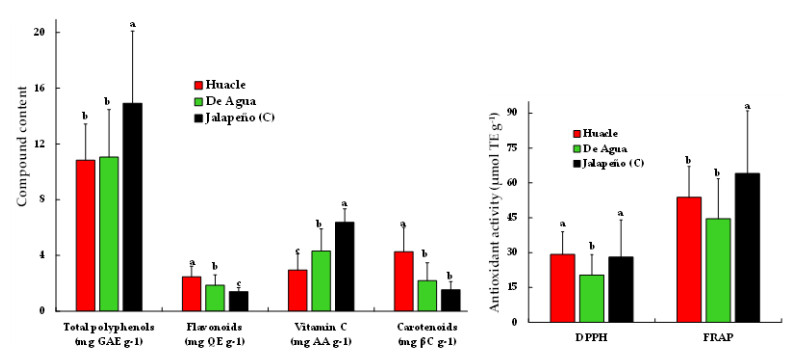
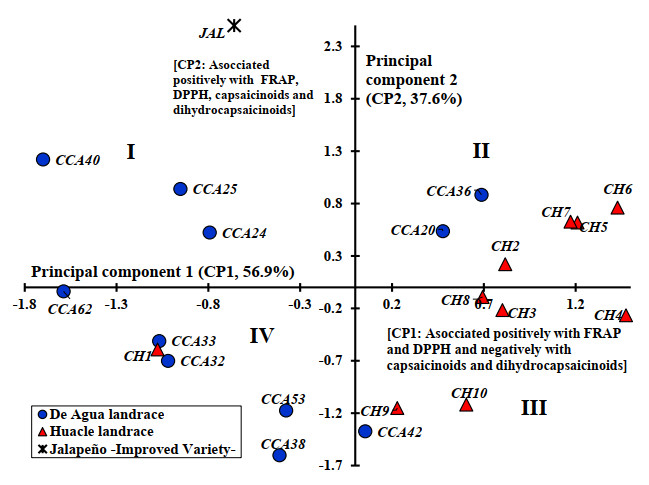
Farmers' varieties or landraces of chili are regularly heterogeneous, selected and preserved by small traditional farmers and highly demanded by regional consumers. The objective of this study was to evaluate the variation in the content of phenolic compounds, vitamin C, carotenoids, capsaicinoids and antioxidant activity in fruits of a population collection of the landraces Huacle and De Agua, which originated in Oaxaca, Mexico, and a commercial variety of Jalapeño (control). The collection was grown in greenhouse conditions under a random block design. At harvest, a sample of ripe fruits was obtained to evaluate the content of phenolic compounds, vitamin C and antioxidant activity by UV–visible spectrophotometry and the concentration of capsaicin and dihydrocapsaicin was measured by high-resolution liquid chromatography. Significant differences were observed between the Huacle and De Agua landraces and between these and Jalapeño. The studied fruits exhibit the following pattern for flavonoid and carotenoid contents: Huacle > De Agua > Jalapeño. The opposite pattern was observed for total polyphenol and vitamin C contents: Jalapeño > De Agua > Huacle. The general pattern for capsaicinoids in fruits was Jalapeño > De Agua > Huacle. Huacle and De Agua populations showed high variability in all compounds evaluated, with positive correlations with antioxidant activity. The capsaicin content in Huacle populations varied ranging from 7.4 to 26.2 mg 100 g-1 and De Agua ranged from 12.4 to 46.8 mg 100 g-1.
Citation: Rosalía García-Vásquez, Araceli Minerva Vera-Guzmán, José Cruz Carrillo-Rodríguez, Mónica Lilian Pérez-Ochoa, Elia Nora Aquino-Bolaños, Jimena Esther Alba-Jiménez, José Luis Chávez-Servia. Bioactive and nutritional compounds in fruits of pepper (Capsicum annuum L.) landraces conserved among indigenous communities from Mexico[J]. AIMS Agriculture and Food, 2023, 8(3): 832-850. doi: 10.3934/agrfood.2023044
Farmers' varieties or landraces of chili are regularly heterogeneous, selected and preserved by small traditional farmers and highly demanded by regional consumers. The objective of this study was to evaluate the variation in the content of phenolic compounds, vitamin C, carotenoids, capsaicinoids and antioxidant activity in fruits of a population collection of the landraces Huacle and De Agua, which originated in Oaxaca, Mexico, and a commercial variety of Jalapeño (control). The collection was grown in greenhouse conditions under a random block design. At harvest, a sample of ripe fruits was obtained to evaluate the content of phenolic compounds, vitamin C and antioxidant activity by UV–visible spectrophotometry and the concentration of capsaicin and dihydrocapsaicin was measured by high-resolution liquid chromatography. Significant differences were observed between the Huacle and De Agua landraces and between these and Jalapeño. The studied fruits exhibit the following pattern for flavonoid and carotenoid contents: Huacle > De Agua > Jalapeño. The opposite pattern was observed for total polyphenol and vitamin C contents: Jalapeño > De Agua > Huacle. The general pattern for capsaicinoids in fruits was Jalapeño > De Agua > Huacle. Huacle and De Agua populations showed high variability in all compounds evaluated, with positive correlations with antioxidant activity. The capsaicin content in Huacle populations varied ranging from 7.4 to 26.2 mg 100 g-1 and De Agua ranged from 12.4 to 46.8 mg 100 g-1.

| [1] |
Krug AS, Drummond EBM, Van Tassel DL, et al. (2023) The next era of crop domestication starts now. Proc Natl Acad Sci 120: e2205769120. https://doi.org/10.1073/pnas.2205769120 doi: 10.1073/pnas.2205769120

|
| [2] | FAOSTAT (2022) Crop and livestock statistics 2020 and 2021. Food and Agriculture Organization of the United Nations (FAO), Rome, Italy. Available from: https://www.fao.org/faostat/en/#data/QCL. |
| [3] |
Karim KMR, Rafii MY, Misran AB, et al. (2021) Current and prospective strategies in the varietal improvement of chilli (Capsicum annuum L.) specially heterosis breeding. Agronomy 11: 2217. https://doi.org/10.3390/agronomy11112217 doi: 10.3390/agronomy11112217
|
| [4] |
Khoury CK, Achicanoy HA, Bjorkman AD, et al. (2016) Origins of food crops connect countries worldwide. Proc R Soc B 283: 20160792. https://dx.doi.org/10.1098/rspb.2016.0792 doi: 10.1098/rspb.2016.0792
|
| [5] |
Perry L, Flannery KV (2007) Precolumbian use of chili peppers in the Valley of Oaxaca, Mexico. Proc Nat Acad Sci 104: 11905–11909. https://doi.org/10.1073/pnas.0704936104 doi: 10.1073/pnas.0704936104
|
| [6] |
Kraft KH, Brown CH, Nabhan GP, et al. (2014) Multiple lines of evidence for the origin of domesticated chili pepper, Capsicum annuum, in Mexico. Proc Natl Acad Sci 111: 6165–6170. https://doi.org/10.1073/pnas.1308933111 doi: 10.1073/pnas.1308933111
|
| [7] |
Taitano N, Bernau V, Jardón-Barbolla L, et al. (2019) Genome-wide genotyping of a novel Mexican chile pepper collection illuminates the history of landraces differentiation after Capsicum annuum L. domestication. Evol Appl 12: 78–92. https://doi.org/10.1111/eva.12651 doi: 10.1111/eva.12651
|
| [8] |
Votova EJ, Baral JB, Bosland PW (2005) Genetic diversity of chile (Capsicum annuum var. annuum L.) landraces from norther New Mexico, Colorado, and Mexico. Econ Bot 59: 8–17. https://doi.org/10.1663/0013-0001(2005)059[0008:GDOCCA]2.0.CO; 2 doi: 10.1663/0013-0001(2005)059[0008:GDOCCA]2.0.CO;2
|
| [9] |
González-Jara P, Moreno-Letelier A, Fraile A, et al. (2011) Impact of human management of the genetic variation of wild pepper, Capsicum annuum var. glabriusculum. PLoS One 6: e28715. https://doi.org/10.1371/journal.pone.0028715 doi: 10.1371/journal.pone.0028715
|
| [10] |
Vera-Guzmán AM, Chávez-Servia JL, Carrillo-Rodríguez JC, et al. (2011) Phytochemical evaluation of wild and cultivated pepper (Capsicum annuum L. and C. pubescens Ruiz & Pav.) from Oaxaca, Mexico. Chil J Agric Res 71: 578–585. http://dx.doi.org/10.4067/S0718-58392011000400013 doi: 10.4067/S0718-58392011000400013
|
| [11] |
Wahyuni Y, Ballester AR, Sudarmonowati E, et al. (2013) Secondary metabolites of Capsicum species and their importance in the human diet. J Nat Prod 76: 783–793. https://doi.org/10.1021/np300898z doi: 10.1021/np300898z
|
| [12] |
Cao S, Chen H, Xiang S, et al. (2015) Anti-cancer effects and mechanisms of capsaicin in chili peppers. Am J. Plant Sci 6: 3075–3081. http://doi.org/10.4236/ajps.2015.619300 doi: 10.4236/ajps.2015.619300
|
| [13] |
Chamikara MDM, Dissanayake DRRP, Ishan M, et al. (2016) Dietary, anticancer and medicinal properties of the phytochemicals in chili pepper (Capsicum spp.). Ceylon J Sci 45: 5–20. http://doi.org/10.4038/cjs.v45i3.7396 doi: 10.4038/cjs.v45i3.7396
|
| [14] |
Mazida MM, Salleh MM, Osman H (2005) Analysis of volatile aroma compounds of fresh chilli (Capsicum annuum) during stages of maturity using solid phase microextraction (SPME). J Food Comp Anal 18: 427–437. https://doi.org/10.1016/j.jfca.2004.02.001 doi: 10.1016/j.jfca.2004.02.001
|
| [15] | Cázares-Sánchez E, Ramírez-Vallejo P, Castillo-González F, et al. (2005) Capsaicinoids and preference of use in different morphotypes of chili peppers (Capsicum annuum L.) of East-Central Yucatan. Agrociencia 39: 627–238. |
| [16] |
Rodríguez-Burruezo A, Kollmannsberger H, González-Mas MC, et al. (2010) HS-SPME comparative analysis of genotypic diversity in the volatile fraction and aroma-contributing compounds of Capsicum fruits from the annuum-chinense-frutescens complex. J Agric Food Chem 58: 4388–4400. https://doi.org/10.1021/jf903931 doi: 10.1021/jf903931t
|
| [17] |
Ghasemnezhad M, Sherafati M, Payvast GA (2011) Variation in phenolic compounds, ascorbic acid and antioxidant activity of five coloured bell pepper (Capsicum annuum) fruits at two different harvest times. J Funct Foods 3: 44–49. https://doi.org/10.1016/j.jff.2011.02.002 doi: 10.1016/j.jff.2011.02.002
|
| [18] |
Arimboor R, Natarajan RB, Menon KR (2015) Red pepper (Capsicum annuum) carotenoids as a source of natural food colors: analysis and stability-a review. J Food Sci Technol 52: 1258–1271. https://doi.org/10.1007/s13197-014-1260-7 doi: 10.1007/s13197-014-1260-7
|
| [19] |
Eggink PM, Maliepaard C, Tikunov Y, et al. (2012) A taste of sweet pepper: Volatile and non-volatile chemical composition of fresh sweet pepper (Capsicum annuum) in relation to sensory evaluation of taste. Food Chem 132: 301–310. https://doi.org/10.1016/j.foodchem.2011.10.081 doi: 10.1016/j.foodchem.2011.10.081
|
| [20] |
Álvarez-Parrilla E, de la Rosa LA, Amarowicz R, et al. (2011) Antioxidant activity of fresh and processed Jalapeño and Serrano peppers. J Agric Food Chem 59: 163–173. https://doi.org/10.1021/jf103434u doi: 10.1021/jf103434u
|
| [21] |
Ornelas-Paz JJ, Martínez-Burrola JM, Ruiz-Cruz S, et al. (2010) Effect of cooking on the capsaicinoids and phenolics contents of Mexican peppers. Food Chem 119: 1619–1625. https://doi.org/10.1016/j.foodchem.2009.09.05 doi: 10.1016/j.foodchem.2009.09.054
|
| [22] |
Hwang IG, Shin YJ, Lee S, et al. (2012) Effects of different cooking methods on the antioxidant properties of red pepper (Capsicum annuum L.). Prev Nutr Food Sci 17: 286–92. https://doi.org/10.3746/pnf.2012.17.4.286 doi: 10.3746/pnf.2012.17.4.286
|
| [23] |
Hamed M, Kalita D, Bartolo ME, et al. (2019) Capsaicinoids, polyphenols, and antioxidant activities of Capsicum annuum: comparative study of the effect of ripening stage and cooking methods. Antioxidants 8: 364. https://doi.org/10.3390/antiox8090364 doi: 10.3390/antiox8090364
|
| [24] |
Thompson RQ, Phinney KW, Sander LC, et al. (2005) Reversed-phase liquid chromatography and argentation chromatography of the minor capsaicinoids. Anal Bioanal Chem 381: 1432–1440. https://doi.org/10.1007/s00216-005-3098-3 doi: 10.1007/s00216-005-3098-3
|
| [25] |
Cisneros-Pineda O, Torres-Tapia LW, Gutiérrez-Pacheco LC, et al. (2007) Capsaicinoids quantification in chili peppers cultivated in the state of Yucatan, Mexico. Food Chem 104: 1755–1760. https://doi.org/10.1016/j.foodchem.2006.10.076 doi: 10.1016/j.foodchem.2006.10.076
|
| [26] |
Wahyuni Y, Ballester AR, Sudarmonowati E, et al. (2011) Metabolite biodiversity in pepper (Capsicum) fruits of thirty-two diverse accessions: Variation in health-related compounds and implications for breeding. Phytochemistry 72: 1358–1370. https://doi.org/10.1016/j.phytochem.2011.03.016 doi: 10.1016/j.phytochem.2011.03.016
|
| [27] |
García-Jiménez FA, Romero-Castillo PA, Reyes-Dorantes A (2018) Presencia de carotenoides en chile ancho y pasilla (Capsicum annuum L.) en muestras de 10 años y recientes. Polibotánica 46: 259–272. https://doi.org/10.18387/polibotanica.46.17 doi: 10.18387/polibotanica.46.17
|
| [28] |
Sánchez-Toledano BI, Cuevas-Reyes V, Kallas Z, et al. (2021) Preferences in 'jalapeño' pepper attributes: A choice study in Mexico. Foods 10: 3111. https://doi.org/10.3390/foods10123111 doi: 10.3390/foods10123111
|
| [29] | AOAC (2005) Association of Official Agricultural Chemists, Ash of flour. 17th, AOAC International Publisher, Gaithersburg, USA. |
| [30] |
Dürüst N, Sümengen D, Dürüst Y (1997) Ascorbic acid and element contents of foods of Trabzon (Turkey). J Agric Food Chem 45: 2085–2087. https://doi.org/10.1021/jf9606159 doi: 10.1021/jf9606159
|
| [31] |
Singleton VL, Rossi JA (1965) Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am J Enol Vitic 16: 144–158. https://doi.org/10.5344/ajev.1965.16.3.144 doi: 10.5344/ajev.1965.16.3.144
|
| [32] |
Lin JY, Tang CY (2007) Determination of total phenolic and flavonoid contents in selected fruits and vegetables, as well as their stimulatory effects on mouse splenocyte proliferation. Food Chem 101: 140–147. https://doi.org/10.1016/j.foodchem.2006.01.014 doi: 10.1016/j.foodchem.2006.01.014
|
| [33] |
Brand-Williams W, Cuvelier ME, Berset CLWT (1995) Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci Technol 28: 25–30. https://doi.org/10.1016/S0023-6438(95)80008-5 doi: 10.1016/S0023-6438(95)80008-5
|
| [34] |
Benzie IF, Strain JJ (1996) The ferric reducing ability of plasma (FRAP) as a measure of "antioxidant power": The FRAP assay. Anal Biochem 239: 70–76. https://doi.org/10.1006/abio.1996.0292 doi: 10.1006/abio.1996.0292
|
| [35] |
Othman ZAA, Ahmed YBH, Habila MA, et al. (2011) Determination of capsaicin and dihydrocapsaicin in Capsicum fruit samples using high performance liquid chromatography. Molecules 16: 8919–8929. https://doi.org/10.3390/molecules16108919 doi: 10.3390/molecules16108919
|
| [36] | Juangsamoot J, Ruangviriyachai C, Techaeongstien S, et al. (2012) Determination of capsaicin and dihydrocapsaicin in some hot chilli varieties by RP-HPLC-PDA after magnetic stirring extraction and clean up with C18 cartridge. Int Food Res J 19: 1217–1226. |
| [37] | SAS Institute Inc. (SAS) (2006) Base SASⓇ 9.1.3 Procedures Guide (2nd edition, Volumes 1–4). SAS Institute Inc.: Cary, NC, USA. |
| [38] | Singh R (2016) Chemotaxonomy: A tool for plant classification. J Med Plants Stud 4: 90–93. |
| [39] |
Hervert-Hernandez D, Sayago-Ayerdi SG, Goñi I (2010). Bioactive compounds of four hot pepper varieties (Capsicum annuum L.), antioxidant capacity, and intestinal bioaccessibility. J Agric Food Chem 58: 3399–3406. https://doig.org/10.1021/jf904220w doi: 10.1021/jf904220w
|
| [40] |
Martínez-Ispizua E, Martínez-Cuenca MR, Marsal JI, et al. (2021) Bioactive compounds and antioxidant capacity of Valencian pepper landraces. Molecules 26: 1031. https://doig.org/10.3390/molecules 26041031 doi: 10.3390/molecules26041031
|
| [41] |
Ionicǎ ME, Nour V, Trandafir I (2017) Bioactive compounds and antioxidant activity of hot pepper fruits at different stages of growth and ripening. J Appl Bot Food Qual 90: 232–237. https://doig.org/10.5073/JABFQ.2017.090.029 doi: 10.5073/JABFQ.2017.090.029
|
| [42] |
Lahbib K, Dabbou S, Bok SE, et al. (2017) Variation of biochemical and antioxidant activity with respect to the part of Capsicum annuum fruit Tunisian autochthonous cultivars. Ind Crop Prod 104: 164–170. https://doi.org/10.1016/j.indcrop.2017.04.037 doi: 10.1016/j.indcrop.2017.04.037
|
| [43] |
Vazquez-Flores AA, Góngora-Pérez O, Olivas-Orduña I, et al. (2020) Phytochemical profile and antioxidant activity of chiltepin chili (Capsicum annuum var. glabriusculum), Sonora, Mexico. J Food Bioact 11: 57–67. https://doi.org/10.31665/JFB.2020.11237 doi: 10.31665/JFB.2020.11237
|
| [44] | Vera-Guzmán AM, Aquino-Bolaños EN, Heredia-García E, et al. (2017) Flavonoid and capsaicinoid contents and consumption of Mexican chili pepper (Capsicum annuum L.) landraces. In: Justino GC (Ed.), Flavonoids-from Biosynthesis to Human Health, London, UK, InTechOpen, 405–437. https://doi.org/10.5772/68076 |
| [45] |
Ribes-Moya AM, Adalid AM, Raigón MD, et al. (2020) Variation in flavonoids in a collection of peppers (Capsicum sp.) under organic and conventional cultivation: effect of the genotype, ripening stage, and growing system. J Sci Food Agric 100: 2208–2223. https://doi.org/10.1002/jsfa.10245 doi: 10.1002/jsfa.10245
|
| [46] |
Antonio AS, Wiedemann LSM, Junior VV (2018) The genus Capsicum: A phytochemical review of bioactive secondary metabolites. RSC Adv 8: 25767–25784. https://doi.org/10.1039/C8RA02067A doi: 10.1039/C8RA02067A
|
| [47] |
Espichán F, Rojas R, Quispe F, et al. (2022) Metabolomic characterization of 5 native Peruvian chili peppers (Capsicum spp.) as a tool for species discrimination. Food Chem 386: 132704. https://doi.org/10.1016/j.foodchem.2022.132704 doi: 10.1016/j.foodchem.2022.132704
|
| [48] |
Castillo-Velarde ER (2019) Vitamin C in health and disease. Rev Fac Med Hum 19: 95–100. https://doi.org/10.25176/RFMH.v19i4.2351 doi: 10.25176/RFMH.v19i4.2351
|
| [49] |
Rosa-Martínez E, García-Martínez MD, Adalid-Martínez AM, et al. (2021) Fruit composition profile of pepper, tomato and eggplant varieties grown under uniform conditions. Food Res Int 147: 110531. https://doi.org/10.1016/j.foores.2021.110531 doi: 10.1016/j.foodres.2021.110531
|
| [50] |
Agostini-Costa ST, da-Silva-Gomes I, de-Melo LAMP, et al. (2017) Carotenoid and total vitamin C content of peppers from selected Brazilian cultivars. J Food Comp Anal 57: 73–79. https://doi.org/10.1016/j.jfca.2016.12.020 doi: 10.1016/j.jfca.2016.12.020
|
| [51] |
Topuz A, Ozdemir F (2007) Assessment of carotenoids, capsaicinoids and ascorbic acid composition of some selected pepper cultivars (Capsicum annuum L.) grown in Turkey. J Food Comp Anal 20: 596–602. https://doi.org/10.1016/j.jfca.2007.03.007 doi: 10.1016/j.jfca.2007.03.007
|
| [52] |
Kim JS, Ahn J, Lee SJ, et al. (2011) Phytochemicals and antioxidant activity of fruits and leaves of paprika (Capsicum annuum L., var. Special) cultivated in Korea. J Food Sci 76: C193–C198. https://doi.org/10.1111/j.1750-3841.2010.01891.x doi: 10.1111/j.1750-3841.2010.01891.x
|
| [53] |
Deepa N, Kaur C, George B, et al. (2007) Antioxidant constituents in some sweet pepper (Capsicum annuum L.) genotypes during maturity. LWT-Food Sci Technol 40: 121–129. https://doi.org/10.1016/j.lwt.2005.09.016 doi: 10.1016/j.lwt.2005.09.016
|
| [54] |
Chávez-Mendoza C, Sanchez E, Muñoz-Marquez E, et al. (2015) Bioactive compounds and antioxidant activity in different grafted varieties of bell pepper. Antioxidants 4: 427–446. https://doi.org/10.3390/antiox4020427 doi: 10.3390/antiox4020427
|
| [55] |
Paredes-Andrade NJ, Monteros-Altamirano A, Tapia-Bastidas CG, et al. (2020) Morphological, sensorial and chemical characterization of chilli peppers (Capsicum spp.) from the CATIE genebank. Agronomy 10: 1732. https://doi.org/10.3390/agronomy10111732 doi: 10.3390/agronomy10111732
|
| [56] | Medina-Juárez LÁ, Molina-Quijada DM, Sánchez CLDT, et al. (2012) Antioxidant activity of peppers (Capsicum annuum L.) extracts and characterization of their phenolic constituents. Interciencia 37: 588–593. |
| [57] |
Rice-Evans CA, Miller NJ, Paganga G (1996) Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic Biol Med 20: 933–956. 10.1016/0891-5849(95)02227-9 doi: 10.1016/0891-5849(95)02227-9
|
| [58] | Gardner PT, White TA, McPhail DB, et al. (2000) The relative contributions of vitamin C, carotenoids and phenolics to the antioxidant potential of fruit juices. Food Chem 68: 471–474. |
| [59] |
Materska M, Perucka I (2005) Antioxidant activity of the main phenolic compounds isolated from hot pepper fruit (Capsicum annuum L.). J Agric Food Chem 53: 1750–1756. https://doi.org/10.1021/jf035331k doi: 10.1021/jf035331k
|
| [60] |
Kogure K, Goto S, Nishimura M, et al. (2002) Mechanism of potent antiperoxidative effect of capsaicin. Biochim Biophys Acta 1573: 84–92. https://doi.org/10.1016/s0304-4165(02)00335-5 doi: 10.1016/S0304-4165(02)00335-5
|
| [61] |
Sora GTS, Haminiuk CWI, da Silva MV, et al. (2015) A comparative study of the capsaicinoid and phenolic contents and in vitro antioxidant activities of the peppers of the genus Capsicum: an application of chemometrics. J Food Sci Technol 52: 8086–8094. https://doi.org/10.1007/s13197-015-1935-8 doi: 10.1007/s13197-015-1935-8
|

Figures(3) / Tables(6)

Rosalía García-Vásquez, Araceli Minerva Vera-Guzmán, José Cruz Carrillo-Rodríguez, Mónica Lilian Pérez-Ochoa, Elia Nora Aquino-Bolaños, Jimena Esther Alba-Jiménez, José Luis Chávez-Servia. Bioactive and nutritional compounds in fruits of pepper (Capsicum annuum L.) landraces conserved among indigenous communities from Mexico[J]. AIMS Agriculture and Food, 2023, 8(3): 832-850. doi: 10.3934/agrfood.2023044







 DownLoad:
DownLoad: