This work focuses on the area of food waste from the subjective perspective of the consumers themselves. The key source of data is a questionnaire survey with a sample selection of 3,429 respondents from the Czech Republic, Poland, and Slovakia, which are countries with historical and cultural ties. This survey was orchestrated in 2019 and it aimed to uncover the consumers' stances on this area. For every country involved, the results proved that the most common reason for food waste is that the food spoils when stored. One's stance on food waste is influenced by identifiers such as age, education, economic activity, and perceived income. A significant difference between the surveyed countries can be seen in the fact that Poles have over a 10× greater chance of wasting the smallest amount of food. Descriptive statistics, contingency analysis, and logistic regression were used to analyze the data. It is evident that subsequent research is necessary in this area, ideally with a united methodology for every country in the European Union.
Citation: Naďa Hazuchová, Jana Stávková, Agnieszka Siedlecka, Ľudmila Nagyová. Consumers' stance on food waste in the Czech Republic, Poland, and Slovakia[J]. AIMS Agriculture and Food, 2022, 7(3): 637-658. doi: 10.3934/agrfood.2022040
This work focuses on the area of food waste from the subjective perspective of the consumers themselves. The key source of data is a questionnaire survey with a sample selection of 3,429 respondents from the Czech Republic, Poland, and Slovakia, which are countries with historical and cultural ties. This survey was orchestrated in 2019 and it aimed to uncover the consumers' stances on this area. For every country involved, the results proved that the most common reason for food waste is that the food spoils when stored. One's stance on food waste is influenced by identifiers such as age, education, economic activity, and perceived income. A significant difference between the surveyed countries can be seen in the fact that Poles have over a 10× greater chance of wasting the smallest amount of food. Descriptive statistics, contingency analysis, and logistic regression were used to analyze the data. It is evident that subsequent research is necessary in this area, ideally with a united methodology for every country in the European Union.

| [1] | FAO (2014) Save food: Global initiative on food loss and waste reduction definitional framework of food loss (Working Paper). |
| [2] | Borma A (2017) Food waste–A global problem. SEA-Practical Application of Science, Romanian Foundation for Business Intelligence, Editorial Department 353–362. |
| [3] | Tukker A, Huppes G, Guinée J, et al. (2006) Environmental impact of products (EIPRO): Analysis of the life cycle environmental impacts related to the final consumption of the EU-25. EUROPEAN COMMISSION DIRECTORATE-GENERAL Joint Research Centre. Available from: http://ec.europa.eu/environment/ipp/pdf/eipro_report.pdf. |
| [4] |
Quested TE, Marsh E, Stunell D, et al. (2013) Spaghetti soup: The complex world of food waste behaviours. Resour Conserv Recy 79: 43–51. https://doi.org/10.1016/j.resconrec.2013.04.011 doi: 10.1016/j.resconrec.2013.04.011

|
| [5] |
Grizzetti B, Pretato U, Lassaletta L, et al. (2013) The contribution of food waste to global and European nitrogen pollution. Environ Sci Policy 33: 186–195. https://doi.org/10.1016/j.envsci.2013.05.013 doi: 10.1016/j.envsci.2013.05.013
|
| [6] |
Flanagana A, Priyadarshini A (2021) A study of consumer behaviour towards food-waste in Ireland: Attitudes, quantities and global warming potentials. J Environ Manage 284: 112046. https://doi.org/10.1016/j.jenvman.2021.112046 doi: 10.1016/j.jenvman.2021.112046
|
| [7] |
Martins IBA, Oliveira D, Rosenthal A, et al. (2019) Brazilian consumer's perception of food processing technologies: A case study with fruit juice. Food Res Int 125: 108555. https://doi.org/10.1016/j.foodres.2019.108555 doi: 10.1016/j.foodres.2019.108555
|
| [8] |
Jeswani HK, Figueroa-Torres G, Azapagic A (2021) The extent of food waste generation in the UK and its environmental impacts. Sustain Prod Consump 26: 532–547. https://doi.org/10.1016/j.spc.2020.12.021 doi: 10.1016/j.spc.2020.12.021
|
| [9] | Gustavsson J, Cederberg C, Sonesson U, et al. (2011) Global food Losses and food waste: Extension, causes and prevention. Available from: http://www.fao.org/3/a-i2697e.pdf. |
| [10] |
Porter SD, Reay DS (2016) Addressing food supply chain and consumption inefficiencies: potential for climate change mitigation. Reg Environ Change 16: 2279–2290. https://doi.org/10.1007/s10113-015-0783-4 doi: 10.1007/s10113-015-0783-4
|
| [11] |
Wohner B, Pauer E, Heinrich V, et al. (2019) Packaging-related food losses and waste: An overview of drivers and issues. Sustainability 11: 264. https://doi.org/10.3390/su11010264 doi: 10.3390/su11010264
|
| [12] | Rezaei M, Liu B (2017) Food loss and waste in the food supply chain. International Nut and Dried Fruit Council 26–27. |
| [13] |
Vilarino MV, Franco C, Quarrington C (2017) Food loss and waste reduction as an integral part of a circular economy. Front Environ Sci 5: 21. https://doi.org/10.3389/fenvs.2017.00021 doi: 10.3389/fenvs.2017.00021
|
| [14] | FAO (2011) Global food losses and food waste: Extent, causes and preventions. Available from: https://www.fao.org/3/mb060e/mb060e00.htm. |
| [15] |
Scherhaufer S, Moates G, Hartikainen H, et al. (2018) Environmental impacts of food waste in Europe. Waste Manage 77: 98–113. https://doi.org/10.1016/j.wasman.2018.04.038 doi: 10.1016/j.wasman.2018.04.038
|
| [16] |
Brancoli P, Rousta K, Bolton K (2017) Life cycle assessment of supermarket food waste. Resour Conserv Recy 118: 39–46. https://doi.org/10.1016/j.resconrec.2016.11.024 doi: 10.1016/j.resconrec.2016.11.024
|
| [17] | USDA (2022) Food waste and its links to greenhouse gases and climate change. Available from: https://www.usda.gov/media/blog/2022/01/24/food-waste-and-its-links-greenhouse-gases-and-climate-change. |
| [18] | San-Epifanio LE, De Renobales Scheifler M (2015) Envisioning a future without food waste and food poverty: Societal challenges. Wageningen Academic Publishers. https://doi.org/10.3920/978-90-8686-820-9 |
| [19] |
Galli A, Wiedmann T, Ercin E, et al. (2012). Integrating ecological, carbon and water footprint into a "footprint family" of indicators: definition and role in tracking human pressure on the planet. Ecol Indic 16: 100–112. https://doi.org/10.1016/j.ecolind.2011.06.017 doi: 10.1016/j.ecolind.2011.06.017
|
| [20] |
Hoekstra AY, Mekonnen MM (2012) The water footprint of humanity. PNAS 109: 3232–3237. https://doi.org/10.1073/pnas.1109936109 doi: 10.1073/pnas.1109936109
|
| [21] |
Steen-Olsen K, Weinzettel J, Cranston G, et al. (2012) Carbon, land, and water footprint accounts for the European Union: Consumption, production, and displacements through international trade. Environ Sci Technol 46: 10883–10891. https://doi.org/10.1021/es301949t doi: 10.1021/es301949t
|
| [22] |
Jones CM, Kammen DM (2011) Quantifying carbon footprint reduction opportunities for U.S. households and communities. Environ Sci Technol 45: 4088–4095. https://doi.org/10.1021/es102221h doi: 10.1021/es102221h
|
| [23] |
Ridoutt BG, Pfister S (2010). Reducing humanity's water footprint. Environ Sci Technol 44: 6019–6021. https://doi.org/10.1021/es101907z doi: 10.1021/es101907z
|
| [24] | Audsley E, Brander M, Chatterton JC, et al. (2010) How low can we go? An assessment of greenhouse gas emissions from the UK food system and the scope reduction by 2050. Report for the WWF and Food Climate Research Network. Available from: https://dspace.lib.cranfield.ac.uk/handle/1826/6503. |
| [25] |
Leip A, Billen G, Garnier J, et al. (2015). Impacts of European livestock production: Nitrogen, sulphur, phosphorus and greenhouse gas emissions, land-use, water eutrophication and biodiversity. Environ Res Let 10: 115004. https://doi.org/10.1088/1748-9326/10/11/115004 doi: 10.1088/1748-9326/10/11/115004
|
| [26] |
Lesschen JP, van den Berg M, Westhoek HJ, et al. (2011) Greenhouse gas emission profiles of European livestock sectors. Anim Feed Sci Technol 166–167: 16–28. https://doi.org/10.1016/j.anifeedsci.2011.04.058 doi: 10.1016/j.anifeedsci.2011.04.058
|
| [27] |
Gaiani S, Caldeira S, Adorno V, et al. (2018). Food wasters: Profiling consumers' attitude to waste food in Italy. Waste Manage 72: 17–24. https://doi.org/10.1016/j.wasman.2017.11.012 doi: 10.1016/j.wasman.2017.11.012
|
| [28] |
Visschers VHM, Wickli N, Siegrist M (2016) Sorting out food waste behaviour: A survey on the motivators and barriers of self-reported amounts of food waste in households. J Environ Psychol 45: 66–78. https://doi.org/10.1016/j.jenvp.2015.11.007 doi: 10.1016/j.jenvp.2015.11.007
|
| [29] |
Ogunmoroti A, Liu M, Li MY, et al. (2022) Unraveling the environmental impact of current and future food waste and its management in Chinese provinces. Res Environ Sustainability 9: 100064. https://doi.org/10.1016/j.resenv.2022.100064 doi: 10.1016/j.resenv.2022.100064
|
| [30] | Xue L, Liu G (2019) Introduction to global food losses and food waste, In: Saving Food: Production, supply chain, food waste, and food consumption, London: Elsevier Academic Press, 1–31. https://doi.org/10.1016/b978-0-12-815357-4.00001-8 |
| [31] | United Nation, Department of economic and social affairs, sustainable development (2022) Transforming our world: The 2030 Agenda for sustainable development. Available from: https://sdgs.un.org/2030agenda. |
| [32] |
Kummu M, de Moel H, Porkka M, et al. (2012) Lost food, wasted resources: Global food supply chain losses and their impacts on freshwater, cropland, and fertiliser use. Sci Total Environ 438: 477–489. https://doi.org/10.1016/j.scitotenv.2012.08.092 doi: 10.1016/j.scitotenv.2012.08.092
|
| [33] | FAO (2022) Sustainable development goals. Available from: https://www.fao.org/sustainable-development-goals/indicators/1231/en/. |
| [34] |
Kafa N, Jaegler A (2021) Food losses and waste quantification in supply chains: A systematic literature review. British Food J 123: 3502–3521. https://doi.org/10.1108/BFJ-09-2020-0879 doi: 10.1108/BFJ-09-2020-0879
|
| [35] |
de Oliveira MM, Lago A, Dal' Magro GP (2021) Food loss and waste in the context of the circular economy: a systematic review. J Clean Prod 294: 126284. https://doi.org/10.1016/j.jclepro.2021.126284 doi: 10.1016/j.jclepro.2021.126284
|
| [36] | FAO (2019) Understanding the issue before taking action—Food loss and waste: what's the difference? Available from: https://www.fao.org/state-of-food-agriculture/2019/en/. |
| [37] | Buzby JC, Wells FH, Hyman J (2014) The estimated amount, value, and calories of postharvest food losses at the retail and consumer levels in the United States. USDA. Available from: https://www.ers.usda.gov/publications/pub-details/?pubid=43836https: //www.ers.usda.gov/publications/pub-details/?pubid = 43836. |
| [38] |
Chang HH (2021) Is it unethical to waste food? Exploring consumer's ethical perspectives and waste intentions. Curr Psychol 1–15. https://doi.org/10.1007/s12144-020-01257-3 doi: 10.1007/s12144-020-01257-3
|
| [39] |
Lebersorger S, Schneider F (2011) Discussion on the methodology for determining food waste in household waste composition studies. Waste Manage 31: 1924–1933. https://doi.org/10.1016/j.wasman.2011.05.023 doi: 10.1016/j.wasman.2011.05.023
|
| [40] |
Parizeau K, Von Massow M, Martin R (2015) Household-level dynamics of food waste production and related beliefs, attitudes, and behaviours in Guelph, Ontario. Waste Manage 35: 207–217. https://doi.org/10.1016/j.wasman.2014.09.019 doi: 10.1016/j.wasman.2014.09.019
|
| [41] |
Williams H, Wikström F, Otterbring T, et al. (2012) Reasons for household food waste with special attention to packaging. J Clean Prod 24: 141–148. https://doi.org/10.1016/j.jclepro.2011.11.044 doi: 10.1016/j.jclepro.2011.11.044
|
| [42] |
Graham-Rowe E, Jessop DC, Sparks P (2019) Self-affirmation theory and pro-environmental behaviour: Promoting a reduction in household food waste. J Environ Psychol 62: 124–132. https://doi.org/10.1016/j.jenvp.2019.02.003 doi: 10.1016/j.jenvp.2019.02.003
|
| [43] |
Stefan V, van Herpen E, Tudoran AA, et al. (2013) Avoiding food waste by Romanian consumers: The importance of planning and shopping routines. Food Qual Prefer 28: 375–381. https://doi.org/10.1016/j.foodqual.2012.11.001 doi: 10.1016/j.foodqual.2012.11.001
|
| [44] | Ehrlich PR, Holdren JP (1994) Population increase causes increased environmental damage, In: Environmental ethics: Reading in theory and application, Boston, London: Jones and Barlett Publishers. |
| [45] |
Sanne C (2002) Willing consumers—or locked-in? Policies for sustainable consumption. Ecol Econ 42: 273–287. https://doi.org/10.1016/S0921-8009(02)00086-1 doi: 10.1016/S0921-8009(02)00086-1
|
| [46] |
Røpke I (1999) The dynamics of willingness to consume. Ecol Econ 28: 399–420. https://doi.org/10.1016/S0921-8009(98)00107-4 doi: 10.1016/S0921-8009(98)00107-4
|
| [47] | Vieira VA (2008) Consumer behavior and managerial decision making. RAC 12: 253–255. |
| [48] | Fischer ARH (2015) Consumer behavior and food science, In: Reference module in food science, Elsevier. https://doi.org/10.1016/B978-0-08-100596-5.03335-7 |
| [49] |
Ananda J, Karunasena GG, Mitsis A, et al. (2021) Analysing behavioural and socio-demographic factors and practices influencing Australian household food waste. J Clean Prod 306: 127280. https://doi.org/10.1016/j.jclepro.2021.127280 doi: 10.1016/j.jclepro.2021.127280
|
| [50] |
Gatley A, Caraher M, Lang T (2014) A qualitative, cross cultural examination of attitudes and behaviour in relation to cooking habits in France and Britain. Appetite 75: 71–81. https://doi.org/10.1016/j.appet.2013.12.014 doi: 10.1016/j.appet.2013.12.014
|
| [51] | Bloom J (2010) American wasteland: How America throws away nearly half of its food (and what we can do about it), Cambridge, MA: Da Capo Press. |
| [52] | Menzel P, D'aluisio F (2008) What the world eats, Berkeley, California: Tricycle Press. |
| [53] |
Di Talia E, Simeone M, Scarpato D (2019) Consumer behaviour types in household food waste. J Clean Prod 214: 166–172. https://doi.org/10.1016/j.jclepro.2018.12.216 doi: 10.1016/j.jclepro.2018.12.216
|
| [54] |
Mou J, Benyoucef M (2021) Consumer behavior in social commerce: Results from a meta-analysis. Technol Forecast Soc Change 167: 120734. https://doi.org/10.1016/j.techfore.2021.120734 doi: 10.1016/j.techfore.2021.120734
|
| [55] |
Yadav PG, Rai J (2017) The generation Z and their social media usage: A review and a research outline. Glob J Enterprise Inform Syst 9: 110–116. https://doi.org/10.18311/gjeis/2017/15748 doi: 10.18311/gjeis/2017/15748
|
| [56] |
Hazuchová N, Antošová I, Stávková J (2020) Food wastage as a display of consumer behaviour. J Competitiveness 12: 51–66. https://doi.org/10.7441/joc.2020.02.04 doi: 10.7441/joc.2020.02.04
|
| [57] |
Jungowska J, Kulczyński B, Sidor A, et al. (2021) Assessment of factors affecting the amount of food waste in households run by Polish women aware of well-being. Sustainability 13: 976. https://doi.org/10.3390/su13020976 doi: 10.3390/su13020976
|
| [58] | Farr-Wharton G, Foth M, Choi J (2014) Identifying factors that promote consumer behaviours causing expired domestic food waste. J Consum Behav 13: 393–402. |
| [59] | Eurostat (2021) EU statistics on income and living conditions microdata 2004–2019, release 1 in 2021, Available from: https://doi.org/10.2907/EUSILC2004-2019V.2. |
| [60] | Eurostat (2021) Mean consumption expenditure by type of household. Available from: https://ec.europa.eu/eurostat/databrowser/view/hbs_exp_t134/default/table?lang=en. |
| [61] |
Setti M, Falasconi L, Segrè A, et al. (2016) Italian consumers' income and food waste behavior. British Food J 118: 1731–1746. https://doi.org/10.1108/BFJ-11-2015-0427 doi: 10.1108/BFJ-11-2015-0427
|
| [62] |
Ghinea C, Ghiuta OA (2019) Household food waste generation: young consumers behaviour, habits and attitudes. Int J Environ Sci Technol 16: 2185–2200. https://doi.org/10.1007/s13762-018-1853-1 doi: 10.1007/s13762-018-1853-1
|
| [63] |
Pellegrini G, Sillani S, Gregori M. et al. (2019) Household food waste reduction: Italian consumers' analysis for improving food management. British Food J 121: 1382–1397. https://doi.org/10.1108/bfj-07-2018-0425 doi: 10.1108/bfj-07-2018-0425
|
| [64] |
Rohm H, Oostindjer M, Aschemann-Witzel J, et al. (2017) Consumers in a sustainable food supply chain (COSUS): Understanding consumer behavior to encourage food waste reduction. Foods 6: 104. https://doi.org/10.3390/foods6120104 doi: 10.3390/foods6120104
|
| [65] |
Bogevska Z, Berjan S, El Bilali H, et al. (2021). Exploring food shopping, consumption and waste habits in North Macedonia during the COVID-19 pandemic. Socio-Econ Plan Sci 82: 101150. https://doi.org/10.1016/j.seps.2021.101150 doi: 10.1016/j.seps.2021.101150
|
| [66] |
Tomaszewska M, Bilska B, Kołożyn-Krajewska D (2022) The influence of selected food safety practices of consumers on food waste due to its spoilage. Int J Environ Res Public Health 19: 8144. https://doi.org/10.3390/ijerph19138144 doi: 10.3390/ijerph19138144
|
| [67] |
Silvennoinen K, Katajajuuri JM, Hartikainen H, et al. (2014) Food waste volume and composition in Finnish households. British Food J 116: 1058–1068. https://doi.org/10.1108/BFJ-12-2012-0311 doi: 10.1108/BFJ-12-2012-0311
|
| [68] |
Aschemann-Witzel J, De Hooge I, Amani P, et al. (2015). Consumer-related food waste: Causes and potential for action. Sustainability 7: 6457–6477. https://doi.org/10.3390/su7066457 doi: 10.3390/su7066457
|
| [69] |
Samotyja U (2022) Consumer understanding of the 'best before' and 'use by' dates. SJ GMU 122: 82–92. https://doi.org/10.26408/122.07 doi: 10.26408/122.07
|
| [70] |
Van Boxstael S, Devlieghere F, Berkvens D, et al. (2014) Understanding and attitude regarding the shelf life labels and dates on pre-packed food products by Belgian consumers. Food Control 37: 85–92. https://doi.org/10.1016/j.foodcont.2013.08.043 doi: 10.1016/j.foodcont.2013.08.043
|
| [71] | Leib EB, Rice C, Neff R, et al. (2016) Consumer perceptions of date labels: National survey. Center for Health Law & Policy Innovation. Available from: https://chlpi.org/wp-content/uploads/2013/12/Consumer-Perceptions-on-Date-Labels_May-2016.pdf. |
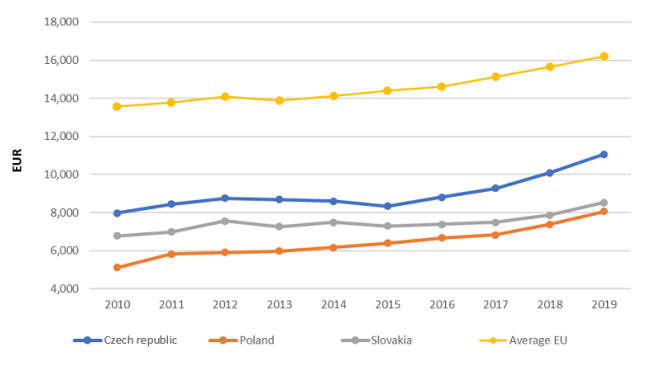
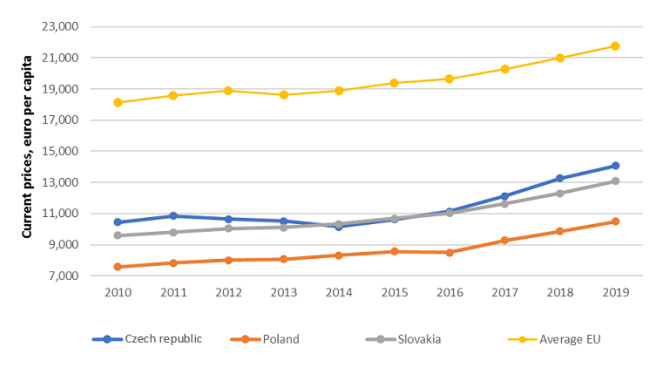
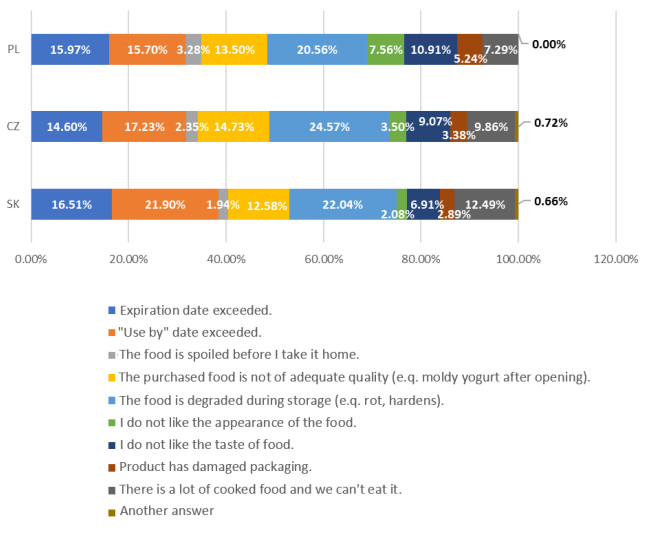
Figures(3) / Tables(9)

Naďa Hazuchová, Jana Stávková, Agnieszka Siedlecka, Ľudmila Nagyová. Consumers' stance on food waste in the Czech Republic, Poland, and Slovakia[J]. AIMS Agriculture and Food, 2022, 7(3): 637-658. doi: 10.3934/agrfood.2022040







 DownLoad:
DownLoad: