Citation: Oleg Sukharev, Ekaterina Voronchikhina. Financial and non-financial investments: comparative econometric analysis of the impact on economic dynamics[J]. Quantitative Finance and Economics, 2020, 4(3): 382-411. doi: 10.3934/QFE.2020018

| [1] |
Afonso A, Aubyn M (2019) Economic growth, public, and private investment returns in 17 OECD economies. Port Econ J 18: 47-65. doi: 10.1007/s10258-018-0143-7

|
| [2] |
Afonso A, Jalles J (2015) How does fiscal policy affect investment? Evidence from a large panel. Int J Financ Econ 20: 310-327. doi: 10.1002/ijfe.1518
|
| [3] |
Allen F, Bartiloro L, Gu X, et al. (2018) Does economic structure determine financial structure? J Int Econ 114: 389-409. doi: 10.1016/j.jinteco.2018.08.004
|
| [4] | Baltagi B (2011) Econometrics, Fifth Edition, Berlin: Springer Cop., 410. |
| [5] |
Bangake C, Eggoh J C (2011) Further evidence on finance-growth causality: A panel data analysis. Econ Syst 35: 176-188. doi: 10.1016/j.ecosys.2010.07.001
|
| [6] |
Beck R, Georgiadis G, Straub R (2014) The finance and growth nexus revisited. Econ Lett 124: 382-385. doi: 10.1016/j.econlet.2014.06.024
|
| [7] |
Bernier M, Plouffe M (2019) Financial innovation, economic growth, and the consequences of macroprudential policies. Res Econ 73: 162-173. doi: 10.1016/j.rie.2019.04.003
|
| [8] |
Botev J, Égert B, Jawadi F (2019) The nonlinear relationship between economic growth and financial development: Evidence from developing, emerging and advanced economies. Int Econ 160: 3-13. doi: 10.1016/j.inteco.2019.06.004
|
| [9] |
Bucci A, Russo A (2019) Finance and long-term growth: Economic modelling and evidence. J Econ Behav Organ 162: 188-192. doi: 10.1016/j.jebo.2019.05.017
|
| [10] |
Chu LK (2020) Financial structure and economic growth nexus revisited. Borsa Istanbul Rev 20: 24-36. doi: 10.1016/j.bir.2019.08.003
|
| [11] |
Combes JL, Kinda T, Ouedraogo R, et al. (2019) Financial flows and economic growth in developing countries. Econ Model 83: 195-209. doi: 10.1016/j.econmod.2019.02.010
|
| [12] |
Ductor L, Grechyna D (2015) Financial development, real sector, and economic growth. Int Rev Econ Financ 37: 393-405. doi: 10.1016/j.iref.2015.01.001
|
| [13] | Fuente-Mella H, Fuentes JLR, Leiva V (2020) Econometric modeling of productivity and technical efficiency in the Chilean manufacturing industry. Comput Ind Eng 139. |
| [14] |
Ghosh A (2017) How does banking sector globalization affect economic growth? Int Rev Econ Financ 48: 83-97. doi: 10.1016/j.iref.2016.11.011
|
| [15] |
Hillebrand E, Pretis F, Proietti T (2020). Econometric models of climate change: Introduction by the guest editors. J Econometrics 214: 1-5. doi: 10.1016/j.jeconom.2019.05.001
|
| [16] |
Hu M, Zhang J, Chao C (2019) Regional financial efficiency and its non-linear effects on economic growth in China. Int Rev Econ Financ 59: 193-206. doi: 10.1016/j.iref.2018.08.019
|
| [17] |
Ibrahim M, Alagidede P (2018) Effect of financial development on economic growth in sub-Saharan Africa. J Policy Model 40: 1104-1125. doi: 10.1016/j.jpolmod.2018.08.001
|
| [18] |
Kavya TB, Shijin S (2020) Economic development, financial development, and income inequality nexus. Borsa Istanbul Rev 20: 80-93. doi: 10.1016/j.bir.2019.12.002
|
| [19] | Mensi W, Hammoudeh S, Tiwari AK, et al. (2020) Impact of Islamic banking development and major macroeconomic variables on economic growth for Islamic countries: Evidence from panel smooth transition models. Econ Syst 44. |
| [20] |
Mollaahmetoğlu E, Akçalı B (2019) The Missing-Link between Financial Development and Economic Growth: Financial Innovation. Proc Comput Sci 158: 696-704. doi: 10.1016/j.procs.2019.09.105
|
| [21] |
Nguyen YN, Brown K, Skully M (2019) Impact of finance on growth: Does it vary with development levels or cyclical conditions? J Policy Model 41: 1195-1209. doi: 10.1016/j.jpolmod.2019.05.006
|
| [22] |
Peia O, Roszbach K (2015) Finance and growth: Time series evidence on causality. J Financ Stab 19: 105-118. doi: 10.1016/j.jfs.2014.11.005
|
| [23] | Pradhan RP, Arvin MB, Hall JH, et al. (2017a) ASEAN economic growth, trade openness and banking-sector depth. Nexus Economia 18: 359-379 |
| [24] |
Pradhan R, Arvin MB, Bahmani S, et al. (2017b) Finance and growth: Evidence from the ARF countries. Q Rev Econ Financ 66: 136-148. doi: 10.1016/j.qref.2017.01.011
|
| [25] | Redmond T, AliNasir M (2020) Role of natural resource abundance, international trade and financial development in the economic development of selected countries. Resour Policy 66. |
| [26] |
Swamy V, Dharani M (2019) The dynamics of finance-growth nexus in advanced economies. Int Rev Econ Financ 64: 122-146. doi: 10.1016/j.iref.2019.06.001
|
| [27] | Sukharev OS (2020a) Economic crisis as a consequence COVID-19 virus attack: risk and damage assessment. Quant Financ Econ 4: 274-293. |
| [28] | Sukharev OS (2020b) Structural analysis of income and risk dynamics in models of economic growth. Quant Financ Econ 4: 1-18. |
| [29] |
Sukharev OS (2019) The restructuring of the investment portfolio: the risk and effect of the emergence of new combinations. Quant Financ Econ 3: 390-411. doi: 10.3934/QFE.2019.2.390
|
| [30] |
Tongurai J, Vithessonthi C (2018) The impact of the banking sector on economic structure and growth. Int Rev Financ Anal 56: 193-207. doi: 10.1016/j.irfa.2018.01.002
|
| [31] |
Välilä T (2020) Infrastructure and growth: A survey of macro-econometric research. Struct Change Econ Dyn 53: 39-49. doi: 10.1016/j.strueco.2020.01.007
|
| [32] | Wooldridge JM (2016) Introductory Econometrics: A Modern Approach, 6th Edition, South-Western Cengage Learning, 912. |
| [33] | Yıldırım S, Gedikli A, Erdoğan S, et al. (2020) Natural resources rents-financial development nexus: Evidence from sixteen developing countries. Resour Policy 68. |
| [34] |
Zeira J, Zoabi H (2015) Economic growth and sector dynamics. Eur Econ Rev 79: 1-15. doi: 10.1016/j.euroecorev.2015.06.007
|
| [35] |
Zhu X, Asimakopoulos S, Kim J (2020) Financial development and innovation-led growth: Is too much finance better? J Int Money Financ 100: 102083. doi: 10.1016/j.jimonfin.2019.102083
|
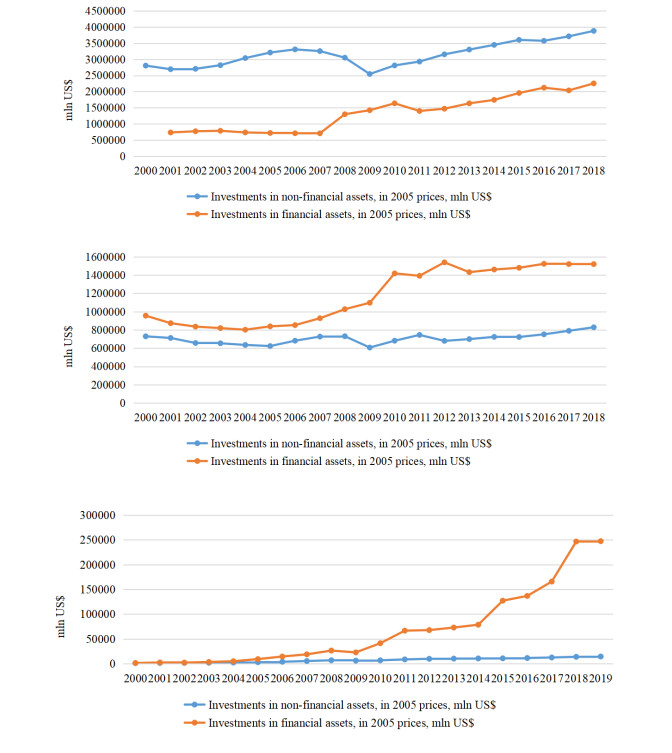
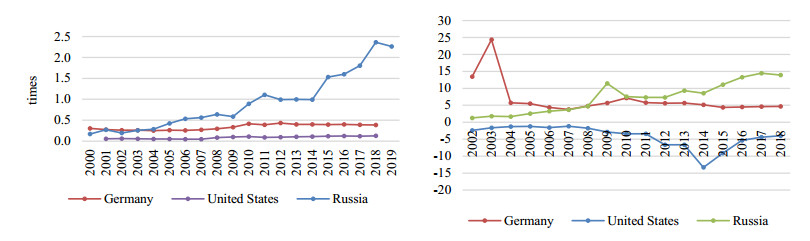
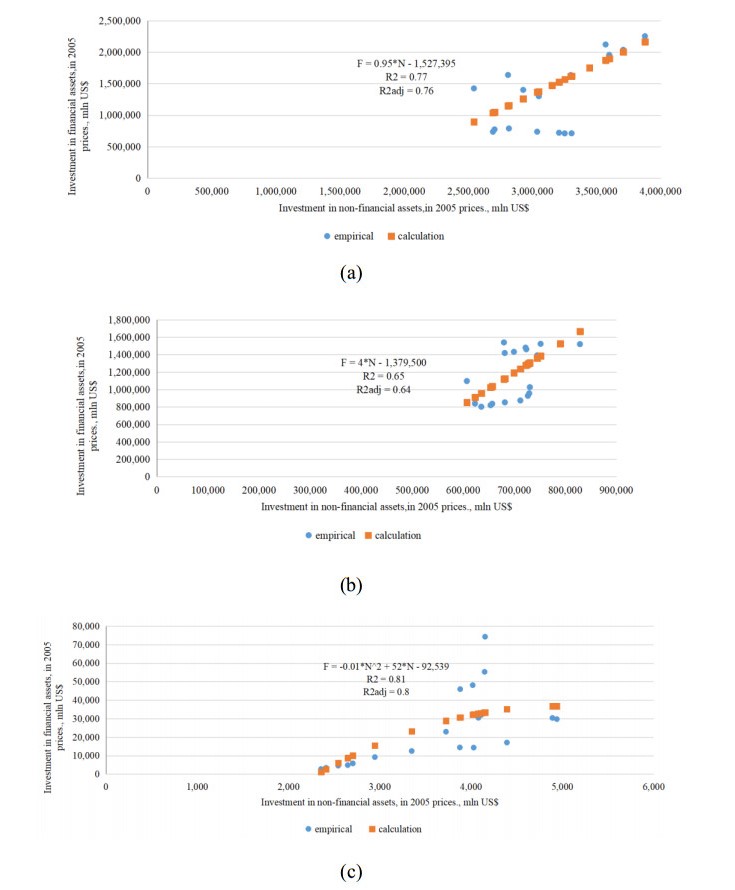
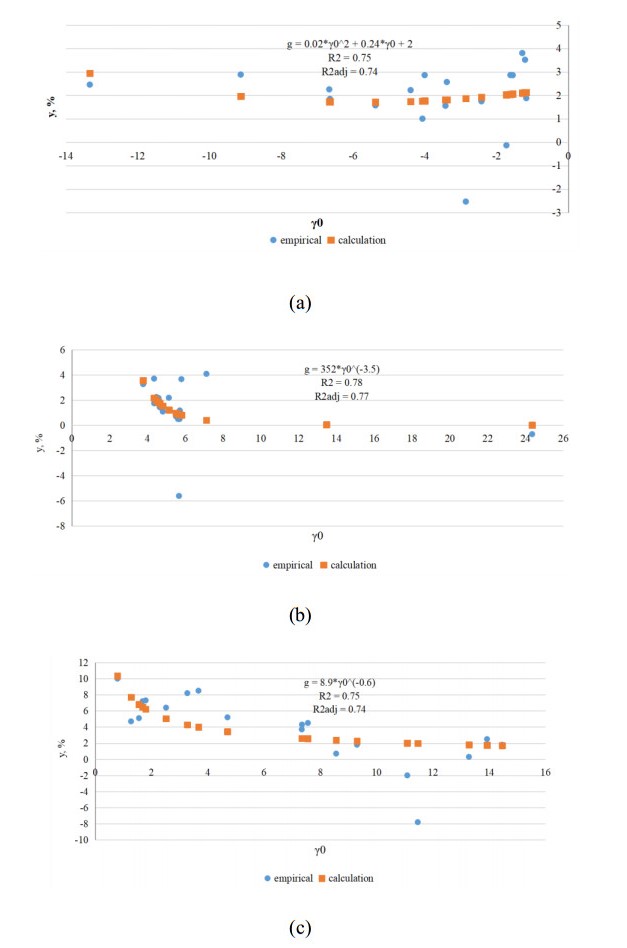
Figures(11) / Tables(13)

Oleg Sukharev, Ekaterina Voronchikhina. Financial and non-financial investments: comparative econometric analysis of the impact on economic dynamics[J]. Quantitative Finance and Economics, 2020, 4(3): 382-411. doi: 10.3934/QFE.2020018







 DownLoad:
DownLoad: