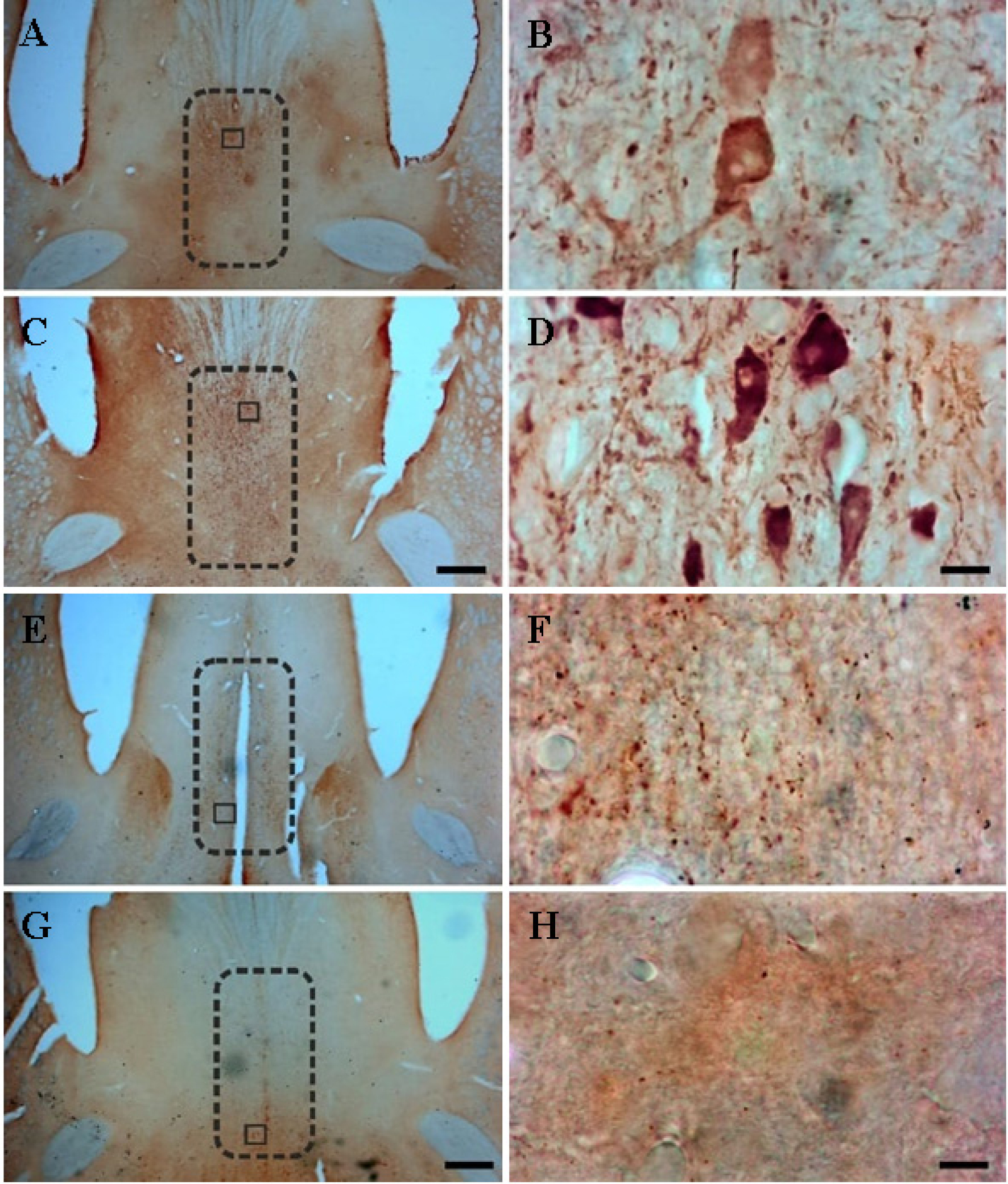
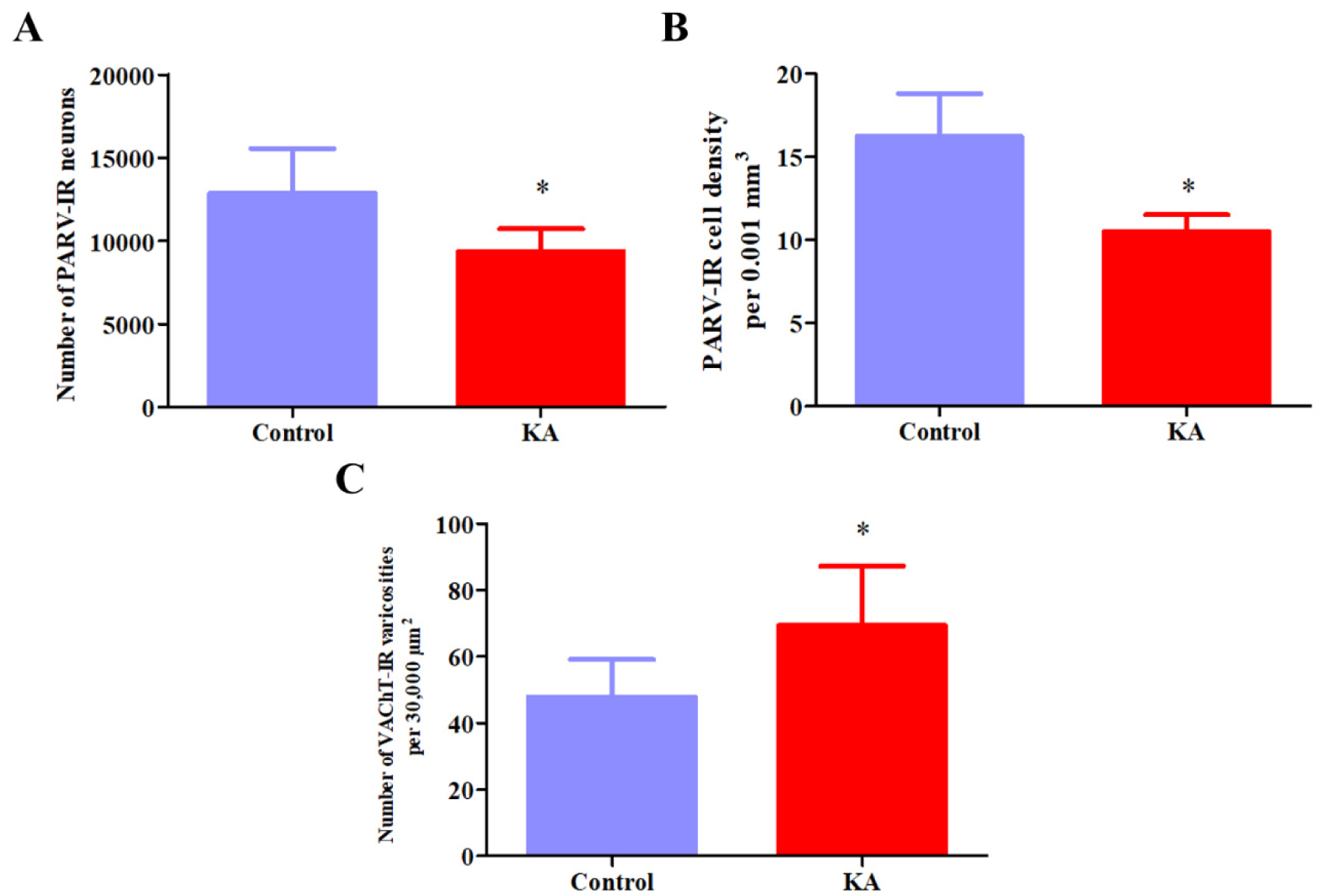
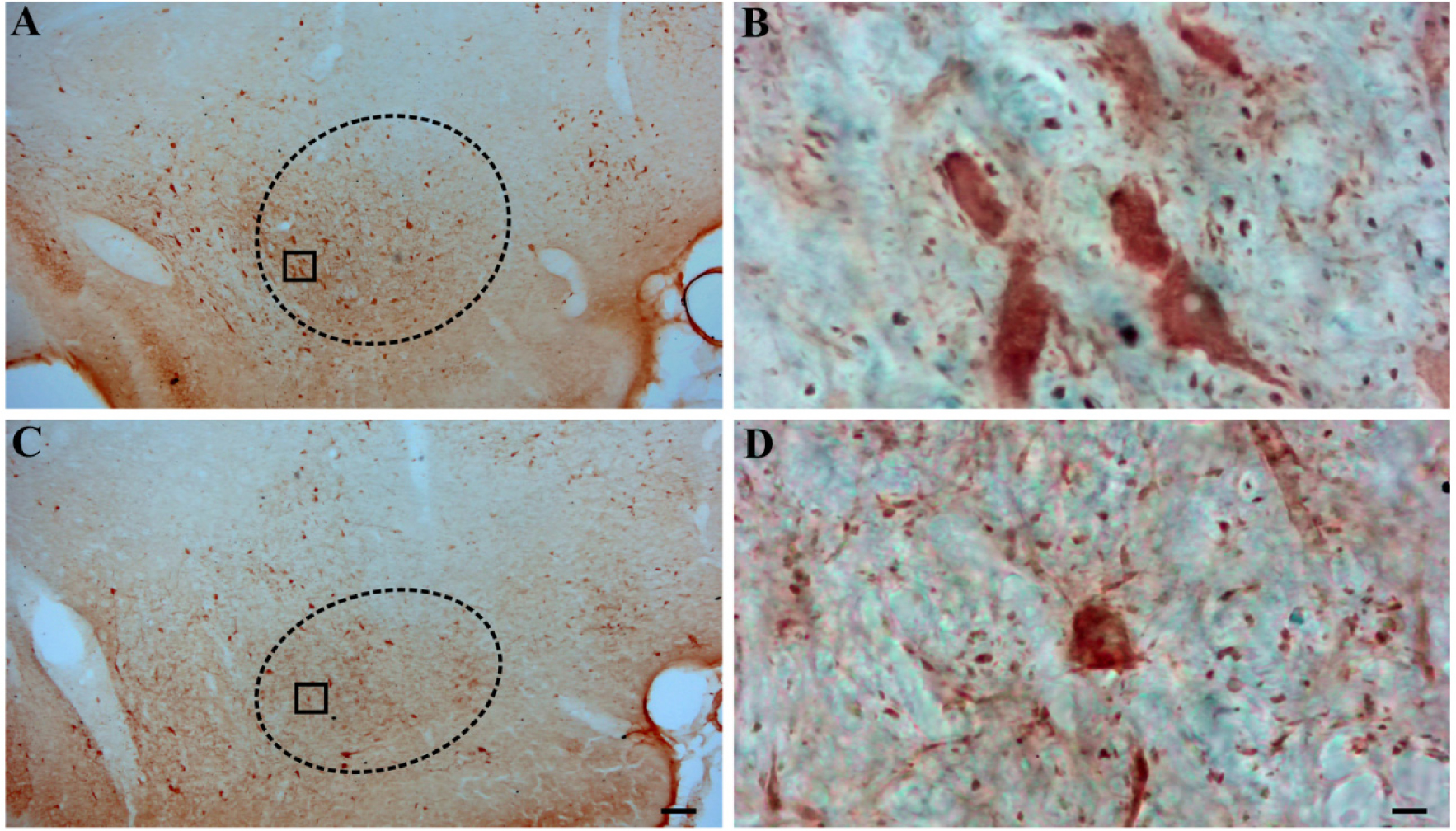
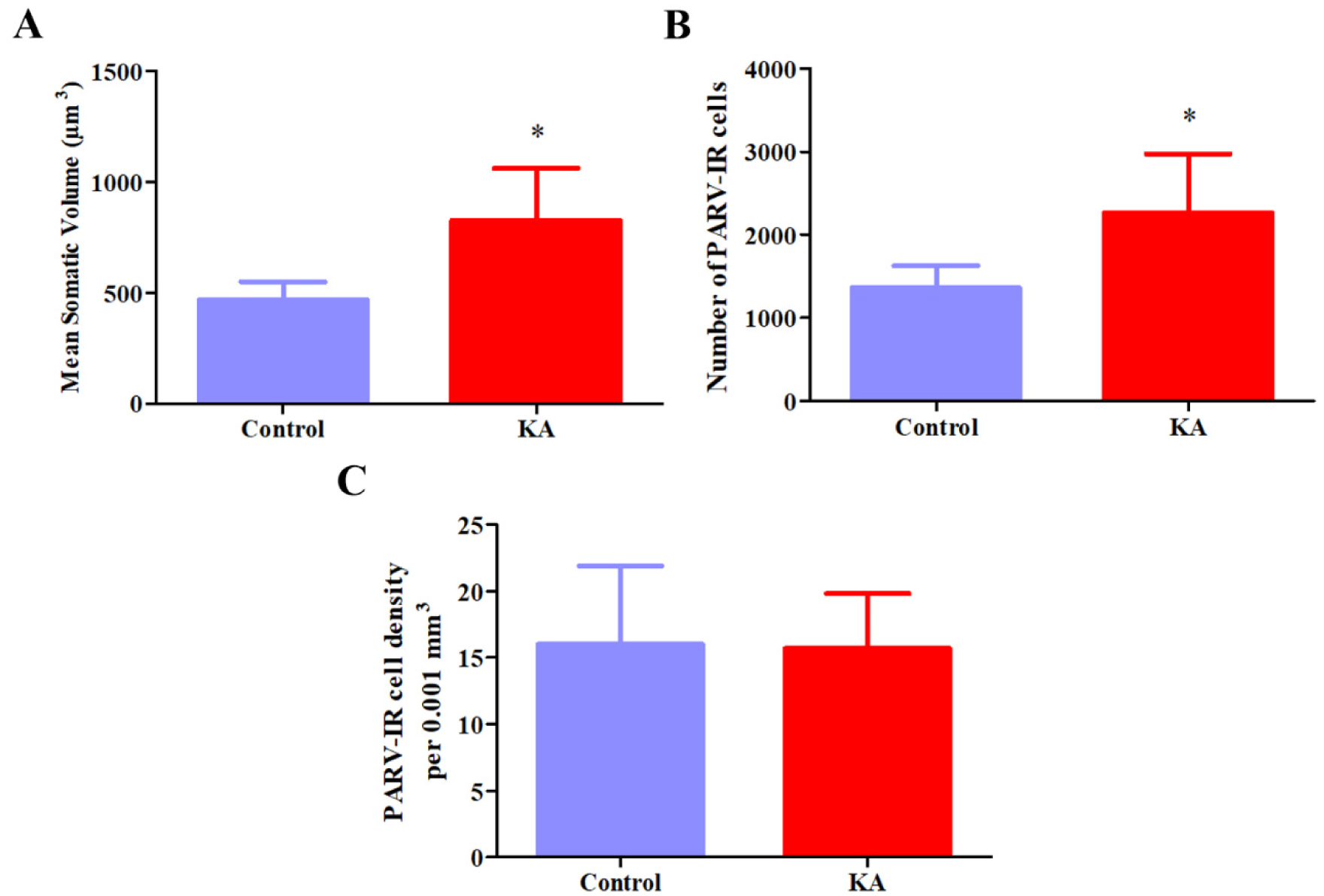
Temporal lobe epilepsy (TLE) is the most prevalent form of epilepsy, through the neuronal mechanisms of this syndrome remain elusive. In addition to the temporal lobe structures, it was found that the basal forebrain cholinergic cells are also involved in epileptogenesis. However, little is known about the involvement of the basal forebrain GABAergic neurons in epilepsy; despite this, they largely project to the temporal lobe and are crucial for the regulation of the hippocampal circuitry. In this study, we assessed epilepsy-induced changes in parvalbumin (PARV) immunoreactive neurons of the medial septum (MS) and of the magnocellular preoptic nucleus (MCPO) using the kainic acid (KA) model in rats. In addition, we estimated the respective changes in the cholinergic varicosities in the MS, where we observed a significant reduction in the PARV cell number (12849 ± 2715 vs. 9372 ± 1336, p = .029) and density (16.2 ± 2.62 vs. 10.5 ± 1.00 per .001 mm3, p =.001), and an increase in the density of cholinergic varicosities (47.9 ± 11.1 vs. 69.4 ± 17.8 per 30,000 µm2, p =.036) in KA-treated animals. In the MCPO, these animals showed a significant increase in somatic volume (827.9 ± 235.2 µm3 vs. 469.9 ± 79.6 µm3, p = .012) and total cell number (2268.6 ± 707.1 vs. 1362.4 ± 262.0, p =.028). These results show that the basal forebrain GABAergic cell populations undergo numerical and morphological changes in epileptic animals, which may contribute to an increased vulnerability of brain circuits to epilepsy and epilepsy-related functional impairments.
Citation: Ruben Carvalho, Alisa N. Lukoyanova, João Casalta-Lopes, Nikolay V. Lukoyanov, Joana Isabel Soares. Plastic rearrangement of basal forebrain parvalbumin-immunoreactive neurons in the kainite model of epilepsy[J]. AIMS Neuroscience, 2023, 10(4): 300-314. doi: 10.3934/Neuroscience.2023023
Temporal lobe epilepsy (TLE) is the most prevalent form of epilepsy, through the neuronal mechanisms of this syndrome remain elusive. In addition to the temporal lobe structures, it was found that the basal forebrain cholinergic cells are also involved in epileptogenesis. However, little is known about the involvement of the basal forebrain GABAergic neurons in epilepsy; despite this, they largely project to the temporal lobe and are crucial for the regulation of the hippocampal circuitry. In this study, we assessed epilepsy-induced changes in parvalbumin (PARV) immunoreactive neurons of the medial septum (MS) and of the magnocellular preoptic nucleus (MCPO) using the kainic acid (KA) model in rats. In addition, we estimated the respective changes in the cholinergic varicosities in the MS, where we observed a significant reduction in the PARV cell number (12849 ± 2715 vs. 9372 ± 1336, p = .029) and density (16.2 ± 2.62 vs. 10.5 ± 1.00 per .001 mm3, p =.001), and an increase in the density of cholinergic varicosities (47.9 ± 11.1 vs. 69.4 ± 17.8 per 30,000 µm2, p =.036) in KA-treated animals. In the MCPO, these animals showed a significant increase in somatic volume (827.9 ± 235.2 µm3 vs. 469.9 ± 79.6 µm3, p = .012) and total cell number (2268.6 ± 707.1 vs. 1362.4 ± 262.0, p =.028). These results show that the basal forebrain GABAergic cell populations undergo numerical and morphological changes in epileptic animals, which may contribute to an increased vulnerability of brain circuits to epilepsy and epilepsy-related functional impairments.
Analysis of variance
Basal forebrain
Dorsal raphe
Hippocampal formation
Kainic acid
Laterodorsal tegmental nucleus
Multivariate analysis of variance
Magnocellular preoptic nucleus
Medial septum nucleus
Mammalian target of the rapamycin signalling pathway
Normal horse serum
Parvalbumin
Parvalbumin-immunoreactive
Phosphate-buffered saline
Pedunculopontine nucleus
Status epilepticus
Temporal lobe epilepsy

| [1] |
Fisher RS, Acevedo C, Arzimanoglou A, et al. (2014) A practical clinical definition of epilepsy. Epilepsia 55: 475-82. https://doi.org/10.1111/epi.12550 
|
| [2] |
Moshe SL, Perucca E, Ryvlin P, et al. (2015) Epilepsy: new advances. Lancet 385: 884-98. https://doi.org/10.1016/S0140-6736(14)60456-6
|
| [3] |
Fisher RS, van Emde Boas W, Blume W, et al. (2005) Epileptic seizures and epilepsy: definitions proposed by the International League Against Epilepsy (ILAE) and the International Bureau for Epilepsy (IBE). Epilepsia 46: 470-2. https://doi.org/10.1111/j.0013-9580.2005.66104.x
|
| [4] |
Baxter MG, Chiba AA (1999) Cognitive functions of the basal forebrain. Curr Opin Neurobiol 9: 178-83. https://doi.org/10.1016/S0959-4388(99)80024-5
|
| [5] |
Everitt BJ, Robbins TW (1997) Central cholinergic systems and cognition. Annu Rev Psychol 48: 649-84. https://doi.org/10.1146/annurev.psych.48.1.649
|
| [6] |
Zaborszky L, Duque A (2003) Sleep-wake mechanisms and basal forebrain circuitry. Front Biosci 8: d1146-69. https://doi.org/10.2741/1112
|
| [7] |
Tramoni-Negre E, Lambert I, Bartolomei F, et al. (2017) Long-term memory deficits in temporal lobe epilepsy. Rev Neurol (Paris) 173: 490-497. https://doi.org/10.1016/j.neurol.2017.06.011
|
| [8] |
Friedman A, Behrens CJ, Heinemann U (2007) Cholinergic dysfunction in temporal lobe epilepsy. Epilepsia 48: 126-30. Erratum in:
|
| [9] |
Risold PY (2004) The Septal Region. The Rat Nervous System . Academic Press 605-632. https://doi.org/10.1016/B978-012547638-6/50021-3
|
| [10] |
Chrobak JJ (2000) Septal orchestration of hippocampal network dynamics. The Behavioral Neuroscience of the Septal Region . New York, NY: Springer 71-91. https://doi.org/10.1007/978-1-4612-1302-4_4
|
| [11] |
Soares JI, Afonso AR, Maia GH, et al. (2018) The pedunculopontine and laterodorsal tegmental nuclei in the kainate model of epilepsy. Neurosci Lett 672: 90-95. https://doi.org/10.1016/j.neulet.2018.02.044
|
| [12] |
Freund TF, Antal M (1988) GABA-containing neurons in the septum control inhibitory interneurons in the hippocampus. Nature 336: 170-3. https://doi.org/10.1038/336170a0
|
| [13] |
Geeraedts LM, Nieuwenhuys R, Veening JG (1990) Medial forebrain bundle of the rat: III. Cytoarchitecture of the rostral (telencephalic) part of the medial forebrain bundle bed nucleus. J Comp Neurol 294: 507-36. https://doi.org/10.1002/cne.902940403
|
| [14] |
Paolini AG, McKenzie JS (1997) Intracellular recording of magnocellular preoptic neuron responses to olfactory brain. Neuroscience 78: 229-42. https://doi.org/10.1016/S0306-4522(96)00566-0
|
| [15] |
Semba K, Reiner PB, McGeer EG, et al. (1988) Brainstem afferents to the magnocellular basal forebrain studied by axonal transport, immunohistochemistry, and electrophysiology in the rat. J Comp Neurol 267: 433-53. https://doi.org/10.1002/cne.902670311
|
| [16] |
Rosal Lustosa Í, Soares JI, Biagini G, et al. (2019) Neuroplasticity in Cholinergic Projections from the Basal Forebrain to the Basolateral Nucleus of the Amygdala in the Kainic Acid Model of Temporal Lobe Epilepsy. Int J Mol Sci 20: 5688. https://doi.org/10.3390/ijms20225688
|
| [17] |
Lothman EW, Collins RC (1981) Kainic acid induced limbic seizures: metabolic, behavioral, electroencephalographic and neuropathological correlates. Brain Res 218: 299-318. https://doi.org/10.1016/0006-8993(81)91308-1
|
| [18] |
Nadler JV (1981) Kainic acid as a tool for the study of temporal lobe epilepsy. Life Sci 29: 2031-42. https://doi.org/10.1016/0024-3205(81)90659-7
|
| [19] |
Soares JI, Valente MC, Andrade PA, et al. (2017) Reorganization of the septohippocampal cholinergic fiber system in experimental epilepsy. J Comp Neurol 525: 2690-2705. https://doi.org/10.1002/cne.24235
|
| [20] |
Maia GH, Soares JI, Almeida SG, et al. (2019) Altered serotonin innervation in the rat epileptic brain. Brain Res Bull 152: 95-106. https://doi.org/10.1016/j.brainresbull.2019.07.009
|
| [21] |
Brashear HR, Zaborszky L, Heimer L (1986) Distribution of GABAergic and cholinergic neurons in the rat diagonal band. Neuroscience 17: 439-51. https://doi.org/10.1016/0306-4522(86)90258-7
|
| [22] |
Meibach RC, Siegel A (1977) Efferent connections of the septal area in the rat: an analysis utilizing retrograde and anterograde transport methods. Brain Res 119: 1-20. https://doi.org/10.1016/0006-8993(77)90088-9
|
| [23] |
Maia GH, Quesado JL, Soares JI, et al. (2014) Loss of hippocampal neurons after kainate treatment correlates with behavioral deficits. PLoS One 9: e84722. https://doi.org/10.1371/journal.pone.0084722
|
| [24] |
Cadete-Leite A, Pereira PA, Madeira MD, et al. (2003) Nerve growth factor prevents cell death and induces hypertrophy of basal forebrain cholinergic neurons in rats withdrawn from prolonged ethanol intake. Neuroscience 119: 1055-69. https://doi.org/10.1016/S0306-4522(03)00205-7
|
| [25] | Paxinos G, Watson C (1998) The rat brain in stereotaxic coordinates. San Diego: Academic Press. |
| [26] |
Gundersen HJ, Jensen EB, Kiêu K, et al. (1999) The efficiency of systematic sampling in stereology--reconsidered. J Microsc 193: 199-211. https://doi.org/10.1046/j.1365-2818.1999.00457.x
|
| [27] |
Garrido Sanabria ER, Castañeda MT, Banuelos C, et al. (2006) Septal GABAergic neurons are selectively vulnerable to pilocarpine-induced status epilepticus and chronic spontaneous seizures. Neuroscience 142: 871-83. https://doi.org/10.1016/j.neuroscience.2006.06.057
|
| [28] |
García-Hernández A, Bland BH, Facelli JC, et al. (2010) Septo-hippocampal networks in chronic epilepsy. Exp Neurol 222: 86-92. https://doi.org/10.1016/j.expneurol.2009.12.010
|
| [29] |
Hristova K, Martinez-Gonzalez C, Watson TC, et al. (2021) Medial septal GABAergic neurons reduce seizure duration upon optogenetic closed-loop stimulation. Brain 144: 1576-1589. https://doi.org/10.1093/brain/awab042
|
| [30] |
Takeuchi Y, Harangozó M, Pedraza L, et al. (2021) Closed-loop stimulation of the medial septum terminates epileptic seizures. Brain 144: 885-908. https://doi.org/10.1093/brain/awaa450
|
| [31] |
Devore S, Pender-Morris N, Dean O, et al. (2016) Basal forebrain dynamics during nonassociative and associative olfactory learning. J Neurophysiol 115: 423-33. https://doi.org/10.1152/jn.00572.2015
|
| [32] |
Zheng Y, Tao S, Liu Y, et al. (2022) Basal Forebrain-Dorsal Hippocampus Cholinergic Circuit Regulates Olfactory Associative Learning. Int J Mol Sci 23: 8472. https://doi.org/10.3390/ijms23158472
|
| [33] |
Zheng Y, Feng S, Zhu X, et al. (2018) Different Subgroups of Cholinergic Neurons in the Basal Forebrain Are Distinctly Innervated by the Olfactory Regions and Activated Differentially in Olfactory Memory Retrieval. Front Neural Circuits 12: 99. https://doi.org/10.3389/fncir.2018.00099
|
| [34] |
Henny P, Jones BE (2006) Vesicular glutamate (VGlut), GABA (VGAT), and acetylcholine (VACht) transporters in basal forebrain axon terminals innervating the lateral hypothalamus. J Comp Neurol 496: 453-67. https://doi.org/10.1002/cne.20928
|
| [35] |
Gritti I, Mainville L, Jones BE (1994) Projections of GABAergic and cholinergic basal forebrain and GABAergic preoptic-anterior hypothalamic neurons to the posterior lateral hypothalamus of the rat. J Comp Neurol 339: 251-68. https://doi.org/10.1002/cne.903390206
|
| [36] |
Saunders A, Granger AJ, Sabatini BL (2015) Corelease of acetylcholine and GABA from cholinergic forebrain neurons. Elife 4: e06412. https://doi.org/10.7554/eLife.06412
|
| [37] |
Takács VT, Cserép C, Schlingloff D, et al. (2018) Co-transmission of acetylcholine and GABA regulates hippocampal states. Nat Commun 9: 2848. https://doi.org/10.1038/s41467-018-05136-1
|
| [38] |
Gritti I, Manns ID, Mainville L, et al. (2003) Parvalbumin, calbindin, or calretinin in cortically projecting and GABAergic, cholinergic, or glutamatergic basal forebrain neurons of the rat. J Comp Neurol 458: 11-31. https://doi.org/10.1002/cne.10505
|
| [39] |
Ma S, Hangya B, Leonard CS, et al. (2018) Dual-transmitter systems regulating arousal, attention, learning and memory. Neurosci Biobehav Rev 85: 21-33. https://doi.org/10.1016/j.neubiorev.2017.07.009
|
| [40] |
Holtzman DM, Lowenstein DH (1995) Selective inhibition of axon outgrowth by antibodies to NGF in a model of temporal lobe epilepsy. J Neurosci 15: 7062-70. https://doi.org/10.1523/JNEUROSCI.15-11-07062.1995
|
| [41] |
Bothwell S, Meredith GE, Phillips J, et al. (2001) Neuronal hypertrophy in the neocortex of patients with temporal lobe epilepsy. J Neurosci 21: 4789-800. https://doi.org/10.1523/JNEUROSCI.21-13-04789.2001
|
| [42] | Houser CR (1992) Morphological changes in the dentate gyrus in human temporal lobe epilepsy. Epilepsy Res Suppl 7: 223-34. |
| [43] |
Zeng LH, Rensing NR, Wong M (2009) The mammalian target of rapamycin signaling pathway mediates epileptogenesis in a model of temporal lobe epilepsy. J Neurosci 29: 6964-72. https://doi.org/10.1523/JNEUROSCI.0066-09.2009
|
Figures(4)

Ruben Carvalho, Alisa N. Lukoyanova, João Casalta-Lopes, Nikolay V. Lukoyanov, Joana Isabel Soares. Plastic rearrangement of basal forebrain parvalbumin-immunoreactive neurons in the kainite model of epilepsy[J]. AIMS Neuroscience, 2023, 10(4): 300-314. doi: 10.3934/Neuroscience.2023023







 DownLoad:
DownLoad: