Citation: Dreaves Hilary A. How Health Impact Assessments (HIAs) Help Us to Select the Public Health Policies Most Likely to Maximise Health Gain, on the Basis of Best Public Health Science[J]. AIMS Public Health, 2016, 3(2): 235-241. doi: 10.3934/publichealth.2016.2.235

| [1] | Gothenburg Consensus paper available at HIA Gateway: Public Health England. 1999. Available from: http://tinyurl.com/o3dbn87. |
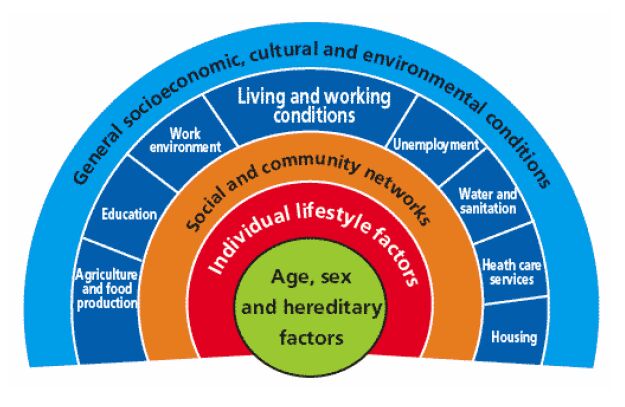
| [2] | Dahlgren,G., Whitehead,M. Socioenvironmental model of health. NHS Scotland webpage. 1991. Available from: http://tinyurl.com/a2leomw. |
| [3] | EU Member States. Declaration on “Health in All Policies”. Rome: December 2007. Available from: http://tinyurl.com/ok4rx5c. |
| [4] | Stone, V (ed.) Health in All Policies. Training Manual (p146). World Health Organisation Geneva. 2015, ISBN 978 92 4 150798 1. Available from: http://tinyurl.com/p7fj2to. |
| [5] | Nowacki, J., Martuzzi, M. Capacity Building in Environment and Health (CBEH) Project. Using impact assessment in environment and health: a framework. Copenhagen: WHO Regional Office for Europe, 2013. Available from: http://tinyurl.com/qcl3dyd. |
| [6] | Gulis,G., Mekel,O (eds). Assessment of Population Health Risks of Policies. 2014, ISBN 978-1-4614-8597-1 (e-book). Available from: http://tinyurl.com/oylehko. |
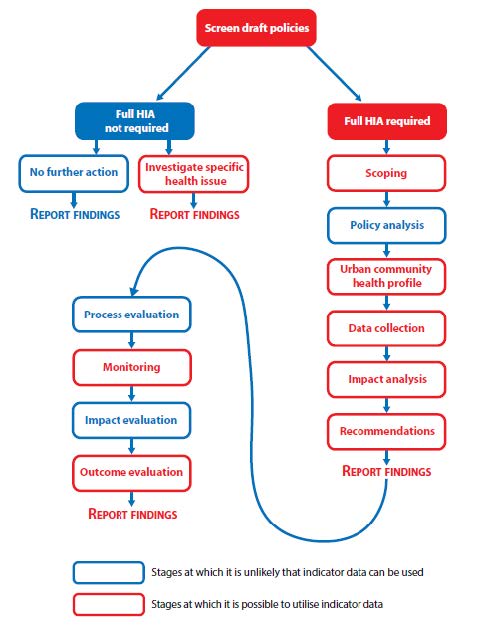
| [7] | Dreaves, H et al. Urban Health Impact Assessment Methodology (UrHIA) IMPACT, University of Liverpool. 2015. Available from: http://tinyurl.com/omun9rz . |
| [8] | Kemm,J. Developing Health Impact Assessment in Wales. National Assembly for Wales. 1999. Available from: http://tinyurl.com/nctlarv. |
| [9] | WHO Regional Office for Europe. Economic cost of the health impact of air pollution in Europe: Clean air, health and wealth. 2015. Available from: http://tinyurl.com/nb7v4dw. |
| [10] | Fehr,R et al. Health in Impact Assessments: Opportunities not to be missed. Copenhagen: WHO Regional Office for Europe. 2014. Available from: http://tinyurl.com/ptlguww. |
| [11] | Climate and Clean Air Coalition. World Health Assembly passes landmark resolution on air pollution and health. 2015. Available from: http://tinyurl.com/q9zmll4. |
| [12] | WHO World Health Assembly. Health and the environment: addressing the health impact of air pollution. Agenda Item 14.6 A68/A/CONF./2 Rev.1 26 May 2015. Available from: http://tinyurl.com/p6w8om4. |
| [13] | World Health Organisation. Sixth report of committee A. Draft A68/75. 27 May 2015. Available from: http://tinyurl.com/pnjtqhe. |
| [14] | Hawe,P et al. A Framework for building capacity to improve health. New South Wales Health Department. 2001, ISBN 0 7347 3124 8. Available from: http://tinyurl.com/paabjzs. |
| [15] | Nowacki, J. Presentation to ESRC Seminar, University of Liverpool, 8th October. 2015. Available from: http://tinyurl.com/q2ksbfj. |
| [16] | Birley M. Health Impact Assessment. Principles and Practice. Routledge (e-book). 2011. Available from: http://tinyurl.com/qdwmshf. |
| [17] | Commission on the Social Determinants of Health (CSDH). Closing the gap in a generation: health equity through action on the social determinants of health. Final Report of the Commission on Social Determinants of Health. Geneva, World Health Organization. 2008. Available from: http://tinyurl.com/oov72qz. |
| [18] | Bell,R., Grobicki, L.,Hamelmann,C. Addressing Social, Economic and Environmental Determinants of Health and the Health Divide in the Context of Sustainable Human Development. UCL Institute of Health Equity for UNDP. 2014. Available from: http://tinyurl.com/negbtyc. |
Figures(2)

Dreaves Hilary A. How Health Impact Assessments (HIAs) Help Us to Select the Public Health Policies Most Likely to Maximise Health Gain, on the Basis of Best Public Health Science[J]. AIMS Public Health, 2016, 3(2): 235-241. doi: 10.3934/publichealth.2016.2.235







 DownLoad:
DownLoad: