Citation: Christy Pu. The Influence of the Common Cold on Self-Rated Health: A Population-Based Study[J]. AIMS Public Health, 2015, 2(3): 247-255. doi: 10.3934/publichealth.2015.3.247

| [1] | Jylha M (2009) What is self-rated health and why does it predict mortality? Towards a unified conceptual model. Soc Sci Med 69: 307-316. |
| [2] |
DeSalvo KB, Fan VS, McDonell MB, et al. (2005) Predicting mortality and healthcare utilization with a single question. Health Serv Res 40: 1234-1246. doi: 10.1111/j.1475-6773.2005.00404.x

|
| [3] |
Pu C, Tang GJ, Huang N, et al. (2011) Predictive power of self-rated health for subsequent mortality risk during old age: analysis of data from a nationally representative survey of elderly adults in Taiwan. J Epidemiol 21: 278-284. doi: 10.2188/jea.JE20100131
|
| [4] |
Goldman N, Glei DA, Chang MC (2004) The role of clinical risk factors in understanding self-rated health. Ann Epidemiol 14: 49-57. doi: 10.1016/S1047-2797(03)00077-2
|
| [5] |
Jylha M, Volpato S, Guralnik JM (2006) Self-rated health showed a graded association with frequently used biomarkers in a large population sample. J Clin Epidemiol 59: 465-471. doi: 10.1016/j.jclinepi.2005.12.004
|
| [6] | Desalvo KB, Muntner P (2011) Discordance between physician and patient self-rated health and all-cause mortality. Ochsner J 11: 232-240. |
| [7] |
Giltay EJ, Vollaard AM, Kromhout D (2012) Self-rated health and physician-rated health as independent predictors of mortality in elderly men. Age Ageing 41: 165-171. doi: 10.1093/ageing/afr161
|
| [8] |
Pietz K, Petersen LA (2007) Comparing self-reported health status and diagnosis-based risk adjustment to predict 1- and 2 to 5-year mortality. Health Serv Res 42: 629-643. doi: 10.1111/j.1475-6773.2006.00622.x
|
| [9] | Sondergaard H, Juul S (2010) Self-rated health and functioning in patients with chronic renal disease. Dan Med Bull 57: A4220. |
| [10] |
Farkas J, Nabb S, Zaletel-Kragelj L, et al. (2009) Self-rated health and mortality in patients with chronic heart failure. Eur J Heart Fail 11: 518-524. doi: 10.1093/eurjhf/hfp038
|
| [11] |
Perruccio AV, Katz JN, Losina E (2012) Health burden in chronic disease: multimorbidity is associated with self-rated health more than medical comorbidity alone. J Clin Epidemiol 65: 100-106. doi: 10.1016/j.jclinepi.2011.04.013
|
| [12] | Galenkamp H, Braam AW, Huisman M, et al. (2012) Seventeen-year time trend in poor self-rated health in older adults: changing contributions of chronic diseases and disability. Eur J Public Health. |
| [13] |
Heller DA, Ahern FM, Pringle KE, et al. (2009) Among older adults, the responsiveness of self-rated health to changes in Charlson comorbidity was moderated by age and baseline comorbidity. J Clin Epidemiol 62: 177-187. doi: 10.1016/j.jclinepi.2008.05.009
|
| [14] |
Charlson ME, Pompei P, Ales KL, et al. (1987) A new method of classifying prognostic comorbidity in longitudinal studies: development and validation. J Chronic Dis 40: 373-383. doi: 10.1016/0021-9681(87)90171-8
|
| [15] | Hoy-Ellis CP, Fredriksen-Goldsen KI (2007) Is AIDS chronic or terminal? The perceptions of persons living with AIDS and their informal support partners. AIDS Care 19: 835-843. |
| [16] | Simon TG, Beland MD, Machan JT, et al. (2012) Charlson Comorbidity Index predicts patient outcome, in cases of inoperable non-small cell lung cancer treated with radiofrequency ablation. Eur J Radiol. |
| [17] |
Unsal A, Resorlu B, Atmaca AF, et al. (2012) Prediction of morbidity and mortality after percutaneous nephrolithotomy by using the Charlson Comorbidity Index. Urology 79: 55-60. doi: 10.1016/j.urology.2011.06.038
|
| [18] | Rattanasompattikul M, Feroze U, Molnar MZ, et al. (2011) Charlson comorbidity score is a strong predictor of mortality in hemodialysis patients. Int Urol Nephrol. |
| [19] |
Koppie TM, Serio AM, Vickers AJ, et al. (2008) Age-adjusted Charlson comorbidity score is associated with treatment decisions and clinical outcomes for patients undergoing radical cystectomy for bladder cancer. Cancer 112: 2384-2392. doi: 10.1002/cncr.23462
|
| [20] |
Charlson ME, Charlson RE, Peterson JC, et al. (2008) The Charlson comorbidity index is adapted to predict costs of chronic disease in primary care patients. J Clin Epidemiol 61: 1234-1240. doi: 10.1016/j.jclinepi.2008.01.006
|
| [21] |
Royston P, Altman DG, Sauerbrei W (2006) Dichotomizing continuous predictors in multiple regression: a bad idea. Stat Med 25: 127-141. doi: 10.1002/sim.2331
|
| [22] |
Royston P, Ambler G, Sauerbrei W (1999) The use of fractional polynomials to model continuous risk variables in epidemiology. Int J Epidemiol 28: 964-974. doi: 10.1093/ije/28.5.964
|
| [23] |
Gunasekara FI, Carter K, Blakely T (2012) Comparing self-rated health and self-assessed change in health in a longitudinal survey: which is more valid? Soc Sci Med 74: 1117-1124. doi: 10.1016/j.socscimed.2011.11.038
|
| [24] |
Giordano GN, Lindstrom M (2010) The impact of changes in different aspects of social capital and material conditions on self-rated health over time: a longitudinal cohort study. Soc Sci Med 70: 700-710. doi: 10.1016/j.socscimed.2009.10.044
|
| [25] |
Nakata A, Takahashi M, Otsuka Y, et al. (2010) Is self-rated health associated with blood immune markers in healthy individuals? Int J Behav Med 17: 234-242. doi: 10.1007/s12529-010-9102-0
|
| [26] |
Idler EL (1993) Age differences in self-assessments of health: age changes, cohort differences, or survivorship? J Gerontol 48: S289-300. doi: 10.1093/geronj/48.6.S289
|
| [27] |
Giron P (2012) Determinants of self-rated health in Spain: differences by age groups for adults. Eur J Public Health 22: 36-40. doi: 10.1093/eurpub/ckq133
|
| [28] |
Calza S, Decarli A, Ferraroni M (2008) Obesity and prevalence of chronic diseases in the 1999-2000 Italian National Health Survey. BMC Public Health 8: 140. doi: 10.1186/1471-2458-8-140
|
| [29] | Lang HC, Lai MS, Chen GT (2004) Outpatient drug expenditures and prescription policies for diseases with high cost to the National Health Insurance system in Taiwan. J Formos Med Assoc 103: 280-285. |
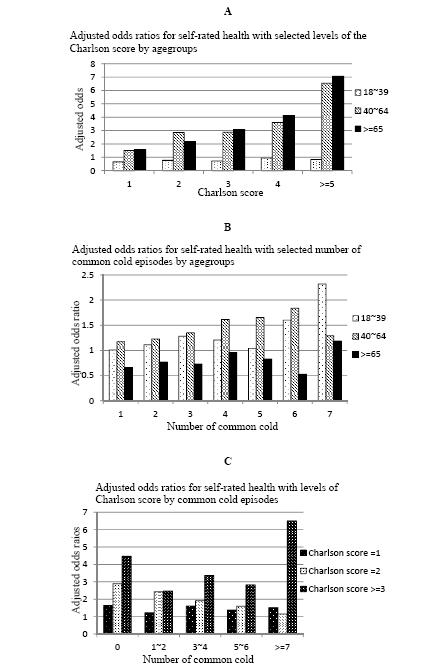
Figures(1) / Tables(1)

Christy Pu. The Influence of the Common Cold on Self-Rated Health: A Population-Based Study[J]. AIMS Public Health, 2015, 2(3): 247-255. doi: 10.3934/publichealth.2015.3.247







 DownLoad:
DownLoad: