




Citation: Guido De Philippis, Filip Rindler. Fine properties of functions of bounded deformation-an approach via linear PDEs[J]. Mathematics in Engineering, 2020, 2(3): 386-422. doi: 10.3934/mine.2020018
| [1] | Eduardo Mercado III . Relating Cortical Wave Dynamics to Learning and Remembering. AIMS Neuroscience, 2014, 1(3): 185-209. doi: 10.3934/Neuroscience.2014.3.185 |
| [2] | Robert A. Moss . A Theory on the Singular Function of the Hippocampus: Facilitating the Binding of New Circuits of Cortical Columns. AIMS Neuroscience, 2016, 3(3): 264-305. doi: 10.3934/Neuroscience.2016.3.264 |
| [3] | Dimitris Pinotsis, Karl Friston . Gamma Oscillations and Neural Field DCMs Can Reveal Cortical Excitability and Microstructure. AIMS Neuroscience, 2014, 1(1): 18-38. doi: 10.3934/Neuroscience.2014.1.18 |
| [4] | Eduardo Mercado III . Learning-Related Synaptic Reconfiguration in Hippocampal Networks: Memory Storage or Waveguide Tuning?. AIMS Neuroscience, 2015, 2(1): 28-34. doi: 10.3934/Neuroscience.2015.1.28 |
| [5] | Robert A. Moss, Jarrod Moss . The Role of Dynamic Columns in Explaining Gamma-band Synchronization and NMDA Receptors in Cognitive Functions. AIMS Neuroscience, 2014, 1(1): 65-88. doi: 10.3934/Neuroscience.2014.1.65 |
| [6] | Ritwik Das, Artur Luczak . Epileptic seizures and link to memory processes. AIMS Neuroscience, 2022, 9(1): 114-127. doi: 10.3934/Neuroscience.2022007 |
| [7] | Ubaid Ansari, Jimmy Wen, Burhaan Syed, Dawnica Nadora, Romteen Sedighi, Denise Nadora, Vincent Chen, Forshing Lui . Analyzing the potential of neuronal pentraxin 2 as a biomarker in neurological disorders: A literature review. AIMS Neuroscience, 2024, 11(4): 505-519. doi: 10.3934/Neuroscience.2024031 |
| [8] | Dai Mitsushima . Contextual Learning Requires Functional Diversity at Excitatory and Inhibitory Synapses onto CA1 Pyramidal Neurons. AIMS Neuroscience, 2015, 2(1): 7-17. doi: 10.3934/Neuroscience.2015.1.7 |
| [9] | Anna Lardone, Marianna Liparoti, Pierpaolo Sorrentino, Roberta Minino, Arianna Polverino, Emahnuel Troisi Lopez, Simona Bonavita, Fabio Lucidi, Giuseppe Sorrentino, Laura Mandolesi . Topological changes of brain network during mindfulness meditation: an exploratory source level magnetoencephalographic study. AIMS Neuroscience, 2022, 9(2): 250-263. doi: 10.3934/Neuroscience.2022013 |
| [10] | Nour Kenaan, Zuheir Alshehabi . A review on recent advances in Alzheimer's disease: The role of synaptic plasticity. AIMS Neuroscience, 2025, 12(2): 75-94. doi: 10.3934/Neuroscience.2025006 |
The role of synapse elimination, observed in humans [1,2,3] and in animals [4,5,6,7,8,9,10], has been generally perceived as the removal, or "pruning", of redundant or weak synapses for the improvement of neural circuit performance. Although structural circuit modification has been suggested in general terms as means for long term memory [11], the specific function and mechanization of synapse elimination have remained essentially unclear. Reports that focal blockade of neurotransmission is more effective in synapse elimination than a whole junction blockade [11], and that synapse elimination precedes axon dismantling [12], have been challenged by claims that synapse elimination is a consequence of whole axon removal [13]. Dynamic firing effects of neural interaction under synapse elimination have been experimentally observed, noting that "active synaptic sites can destabilize inactive synapses in their vicinity" [11], although such effects may involve synapse silencing and reactivation [14] rather than synapse elimination. While early studies have associated synapse elimination with early development [12,15] and childhood [16], others have extended it to puberty [17], and, depending on brain regions, to age 12 for frontal and parietal lobes, to age 16 for the temporal lobe, and to age 20 for the occipital lobe [18]. Yet, grey matter [19] and cognition [20] studies, and persistent evidence of molecular processes involved in synaptic elimination throughout life [21], have suggested its relevance all the way to senescence.
Here, we study the mechanism and the implications of synapse elimination on neural circuit formation, modification and function in a model-based context. Noting that, albeit certain age-related differences in the time constants associated with membrane and synapse plasticity, the firing rate model produces essentially the same dynamic modes [22], we suggest that the model corresponding to maturity and aging represents a viable platform for analyzing the effects of synapse elimination throughout life. We further suggest that synapse weakening and eventual elimination is a consequence of asynchrony in the firing of interacting neurons and circuits. We proceed to show that while whole axon elimination removes asynchronous firing altogether, synapse elimination facilitates interference-free asynchrony between individual neurons and between neural circuit firing, maintaining internal circuit synchrony. This allows for cortical segregation into neurons and circuits having different characteristic firing rate modes corresponding to different cortical functions.
Both the mathematical analysis and the simulation of synapse elimination effects will require some specification of the models involved in order to demonstrate the functional implications. For a single isolated neuron, the firing rate model, evolving from the integrate-and-fire [23] and the conductance-based membrane current[24] paradigms, through cortical averaging [25], neuronal decoding [26] and spiking rate [27] models, is captured, in essence, by the discrete iteration map [28,22].
| υ(k)=αυ(k−1)+βf(ω(k)υ(k−1)+u) | (1) |
where υ(k) is the firing rate, α=exp(−1/τm) and β=1−α, with τmthe membrane time constant, ω(k)is the self-feedback synaptic weight, u is the external membrane activation and
| f(x)={xifx≥00ifx<0 | (2) |
is the conductance-based rectification kernel [29,30,31].
The Bienenstock-Cooper-Munro plasticity rule [32], enhanced by stabilizing modifications [33,34], is a widely recognized, biologically plausible, mathematical representation of the Hebbian learning paradigm [35], taking the discrete form
| ω(k)=εω(k−1)+γ(υ(k−1)−θ(k−1))υ2(k−1) | (3) |
where
| θ(k)=δN∑i=0exp(−i/τθ)υ2(k−i) | (4) |
with ε=exp(−1/τω),γ=1−ε and δ=1/τθ, where τω and τθ are the synaptic and the threshold time constants, respectively.
Different ranges of the time constants, τm, τω and τθ, hypothesized to correspond to different developmental stages (smaller values corresponding to early development), have been shown to produce somewhat different maps of firing rate, possessing, however, essentially similar global attractors [22]. In critical period, immediately following birth and extending into early childhood, the plasticity time constants τω and τθ are assumed to have near-zero values, while the membrane time constant, τm is assume to be higher, producing, by Eq. 1, the map depicted in Figure 1a. The inhibition onset and termination thresholds are h and g, respectively, where the slopes of the map change sign. The dynamic behavior induced by the map depends on the slope at the point p, where the map intersects the diagonal υ(k)=υ(k−1).
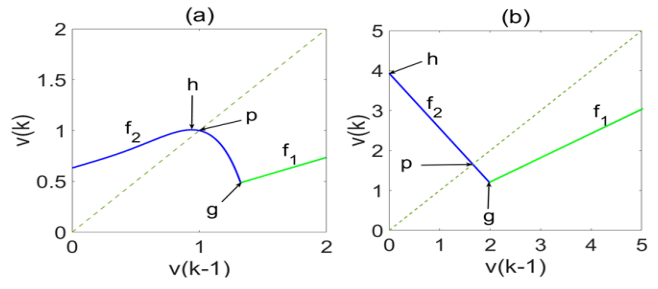 Figure 1. Developmental maps of neuronal firing: (a) critical period, (b) synaptic maturity and rigidity.
Figure 1. Developmental maps of neuronal firing: (a) critical period, (b) synaptic maturity and rigidity.For our present purposes, we continue with the case corresponding to synaptic maturity and rigidity (Figure 1b), so as to stress the relevance of circuit segregation beyond early development. For this case, ω(k) converges to a fixed value ω [34], yielding the model [22]
| υ(k)={f1(υ(k−1))=λ1υ(k−1)forβωυ(k−1)+βu≤0f2(υ(k−1))=λ2υ(k−1)+βuforβωυ(k−1)+βu>0 | (5) |
where
| λ1=α | (6) |
and
| λ2=α+βω | (7) |
and the inhibition onset and offset points become
| h=βu | (8) |
and
| g=βuλ1−λ2 | (9) |
while the fixed point representing the intersection of f2 with the diagonal υ(k)=υ(k−1) is
| p=βu1−λ2 | (10) |
The parameters
| c1=2λ1λ2+1+√1+4λ21 | (11) |
and
| c2=λ1λ2+1 | (12) |
define transition points from one global attractor type of the map to another [28].
Scalar global attractors are graphically described by cobweb diagrams [36,37,38,39], which are initiated at some value, υ(0), on the map, then connected horizontally to the diagonal υ(k)=υ(k−1), then connected vertically to the map, and so on. The cobweb diagrams depicted by the corresponding subplots of Figure 2 represent different global attractor types, which satisfy the following characteristic conditions:
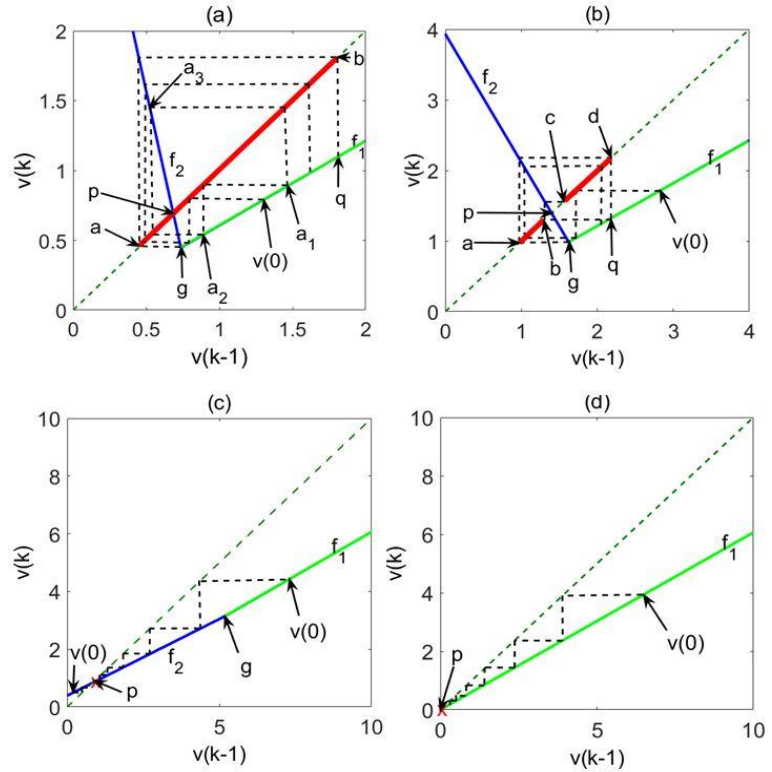 Figure 2. The neuronal global attractor code of firing-rate dynamics: (a) Chaotic,
(b) largely-oscillatory, (c) fixed-point and (d) silent.
Figure 2. The neuronal global attractor code of firing-rate dynamics: (a) Chaotic,
(b) largely-oscillatory, (c) fixed-point and (d) silent.(a) Chaotic attractor. For u>0, λ2<−1, c1≤0, and c2<0, yielding pυ≤qυ (where q is the point obtained from the bend point g by a 4-step cobweb sequence, and where pυ and qυ are the vertical coordinates of the corresponding points on the map in Figure 2a), the attractor is represented in Figure 2a by the interval ab on the diagonal υ(k)=υ(k−1), with a and b created by a cobweb sequence initiating at the bend point g, which defines the boundaries of the attractor, as shown in the figure. An orbit of period three a1→a2→a3→a1, rendering Li-Yorke chaos [40], is defined by a1=f3(a1), a1≠f(a1), where f is the map Eq. 5, and f3(x)=f(f(f(x))). Starting with a1>g, we obtain a2=λ1a1, a3=λ21a1, a1=λ2λ21a1+βu, yielding a1=βu/(1−λ2λ21).
(b) Largely-oscillatory attractor. For u>0, λ2<−1, c1>0, and c2<0, yielding qυ<pυ, the attractor, represented in Figure 2b by the two intervals ab and cd, separated by the repelling interval bc, is largely oscillatory. Within the attractor domain, defined by a cobweb initiating at the bend-point g, trajectories alternate between the two intervals ab and cd. As implied by the cobweb diagram, depending on the circuit parameters, this alternation may, but need not necessarily, repeat precisely the same points, which may then represent oscillatory or cyclically multiplexed dynamics (the two intervals ab and cd then reduce into two or four points, respectively [22,28]).
(c) Fixed-point (constant) attractor. For u>0 and −1<λ2≤1, we have a fixed-point attractor at p. For −1<λ2≤0, the fixed point will be approached by alternate convergence (increasing υ(k) step followed by decreasing υ(k+1) step and vice versa). For 0<λ2≤λ1, convergence will be monotone, bimodal (according to f1 far from p and according to f2 near p, as illustrated by Figure 2c for 0<λ2≤λ1, and unimodal (according to f2) for λ1<λ2≤1.
(d) Silent attractor. For ω=0, the attractor is at the origin, as depicted in Figure 2d.
Specifically, the cobweb diagrams depicted in Figure 2 correspond to the following parameter values:
(a) u=10, τm=2, τω=10000, τθ=0.1, yielding
λ1=0.6055,λ2=−4.7336,c1=−3.1701,c2=−1.8711,ω=−13.5720
(b) u=10,τm=2,τω=10000,τθ=1, yielding
λ1=0.6055,λ2=−1.8160,c1=0.3692,c2=−0.1015,ω=−6.1569
(c) u=1,τm=2,τω=5,τθ=1, yielding λ1=0.6065,λ2=0.5308,ω=−0.1925
(d) u=−1,τm=2,τω=5,τθ=1, yielding λ1=0.6065,λ2=0.6065,ω=0
For each of the cases, the steady-state value of the synaptic weight, ω, was calculated by driving Eqs. 3 and 4, with ω(0)=0 and υ(0)=1, to convergence (practically, this was achieved for N=100), and the corresponding values of λ1,λ2,c1 and c2 were calculated by Eqs. 6, 7, 11 and 12 respectively. As can be verified, the conditions stated above for the attractor types (a) chaotic, (b) largely-oscillatory, (c) fixed-point and (d) silent, are satisfied, respectively, by the parameter values obtained.
For a circuit of n neurons, firing rate and plasticity would be governed by the equations
| υi(k)=αiυi(k−1)+βif(ωiT(k)υ(k−1)+ui) | (13) |
| ωi(k)=εiωi(k−1)+γi[υi(k−1)−θi(k−1)]υ2(k−1) | (14) |
| θi(k)=δiN∑i=0exp(−i/τθi)υi2(k−i) | (15) |
where i=1,2,...,n, υ(k) is the vector of neuronal firing rates, ωi(k) is the vector of input synaptic weights (including self-feedback) corresponding to the i th neuron and υ2 is the vector whose components are the squares of the components of υ. We have used N = 100, yielding convergence of ωi,j,i,j=1,2,…,n, to constant values.
Consider first two neurons, one having the parameters of case (c) of the previous section, hence, a fixed-point global attractor of firing rate, and the other having the parameters of case (b) of the previous section, hence, a largely oscillatory global attractor of firing rate. Figure 3a shows the firing rate sequences of the two neurons, fully connected into a circuit. Eliminating the axon of the first neuron (Figure 3b) reveals the characteristic firing mode of the second neuron alone. Eliminating the axon of the second neuron (Figure 3c) reveals the characteristic firing mode of the first neuron alone. It can be clearly seen by comparison that full connectivity, resulting in the firing sequences displayed by Figure 3a, causes mutual interference between the two neurons. Inter-neuron asynchrony is assumed to imply, by the Hebbian paradigm, weakening of the corresponding synapses [35].
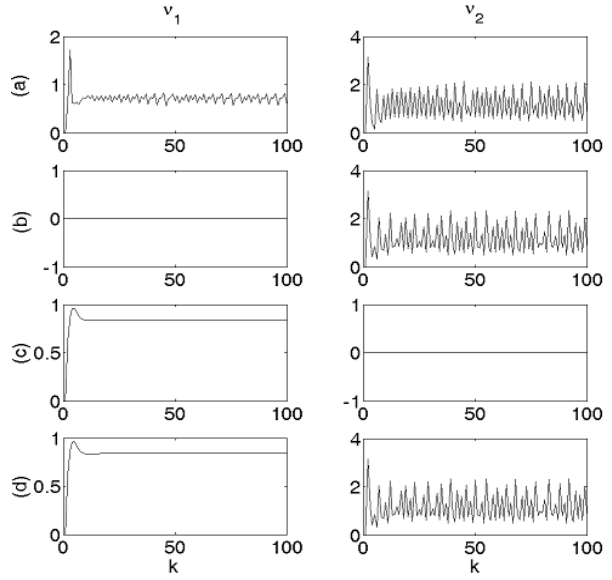 Figure 3. Segregation and interference elimination in a 2-neuron circuit by axon elimination and by synapse elimination.
Figure 3. Segregation and interference elimination in a 2-neuron circuit by axon elimination and by synapse elimination.Eventual elimination of the receiving synapses of both neurons, represented by zero values of the corresponding synaptic weights, allows each of the neurons to display its own characteristic firing-rate mode without interference, as depicted in Figure 3d.
Next consider a fully connected circuit of n identical neurons receiving identical activation. The neurons will fire in synchrony. However, for n > 1, the neuronal firing mode will not be the same as that of an individual isolated neuron with the same property. This can be seen by noting that, for each of the synchronous circuit neurons, Eq. 13 will yield the firing rate model
| υ(k)=αυ(k−1)+βf(nω(k)υ(k−1)+u) | (16) |
which is different from Eq. 1. As N→∞, Eqs. 14 and 15 will take ω(k) to its constant limit value ωf [34], yielding
| υ(k)=αυ(k−1)+βf(nωυ(k−1)+u) | (17) |
which is the firing rate model of an isolated individual neuron with feedback synaptic weight nω. This synchrony equivalence principle allows us to extend our results obtained for the neuronal firing rate modes and for asynchronous neuron segregation to synchronous circuits. Clearly, Eq. 17 will produce a different mode of firing rate dynamics for every circuit size n. This is illustrated by Figure 4, where the four subfigures depict attractors of fully connected circuits having identical neurons, but different circuit sizes. It can be seen that different circuit sizes result in different firing rate modes. Specifically, the circuits represented in Figure 4 obey the model of Eqs.13–15, with N=100 (taking ω(k) to its constant limit ω). For circuits of fully connected identical neurons, having the parameter values u=4, τm=2, τω=300, τθ=0.1 and circuit size values 10, 5, 2 and 1, we obtain the modal parameter and condition values specified below:
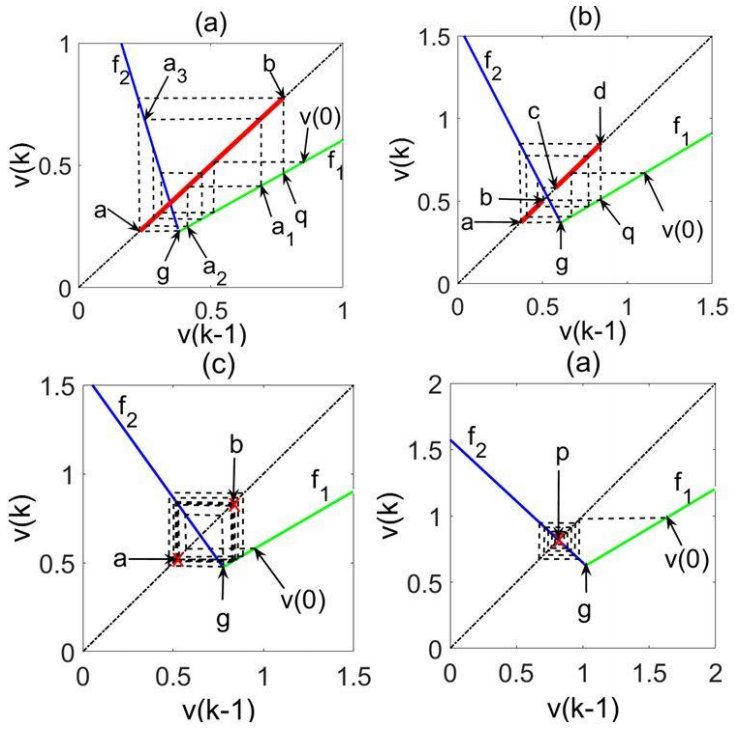 Figure 4. Global attractor types corresponding to fully connected synchronous circuits of (a) ten neurons (chaotic), (b) five neurons (largely-oscillatory), (c) two neurons (oscillatory), and (d) one neuron (fixed point).
Figure 4. Global attractor types corresponding to fully connected synchronous circuits of (a) ten neurons (chaotic), (b) five neurons (largely-oscillatory), (c) two neurons (oscillatory), and (d) one neuron (fixed point).(a) n=10, yielding ω=−0.7870,λ1=0.6065,λ2=−2.4901, c1=−0.4485,c2=−0.5103
(b) n=5, yielding ω=−1.3128,λ1=0.6065,λ2=−1.9762, c1=0.1748,c2=−0.1987
(c) n=2, yielding ω=−2.5628,λ1=0.6065,λ2=−1.4103, c1=0.8614,c2=0.1446
(d) n=1, yielding ω=−3.8993,λ1=0.6065,λ2=−0.9277, c1=1.4467,c2=0.4373
An examination of the conditions for the global attractor types specified in Section 2 shows that the above cases represent (a) chaotic, (b) largely-oscillatory, (c) oscillatory and (d) fixed-point global attractors, as ratified by Figure 4. As, except for the circuit size (i.e., the number of neurons) the neuronal parameters u,τm,τω,τθ are identical for all neurons, the increasing number of neurons in cases (a–d) above may be viewed as representing synchronous circuit segregation into smaller synchronous sub-circuits.
Finally, in order to illustrate the effects of synchronous circuit segregation into synchronous sub-circuits, consider a circuit of three identical neurons with the parameters (identical to those employed in Figure 4) u=4,τm=2,τω=300,τθ=0.1, starting with full connectivity and undergoing modification and segregation by synapse elimination (manifested by setting the corresponding synaptic weight at zero). The changes in circuit connectivity are illustrated in Figure 5, while the resulting changes in the neuronal firing modes, simulated by Eqs. 13–15, are displayed in Figure 6.
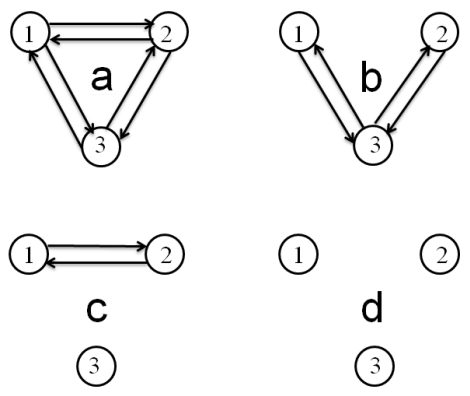 Figure 5. Three-neuron circuit modification and segregation by synapse elimination.
Figure 5. Three-neuron circuit modification and segregation by synapse elimination.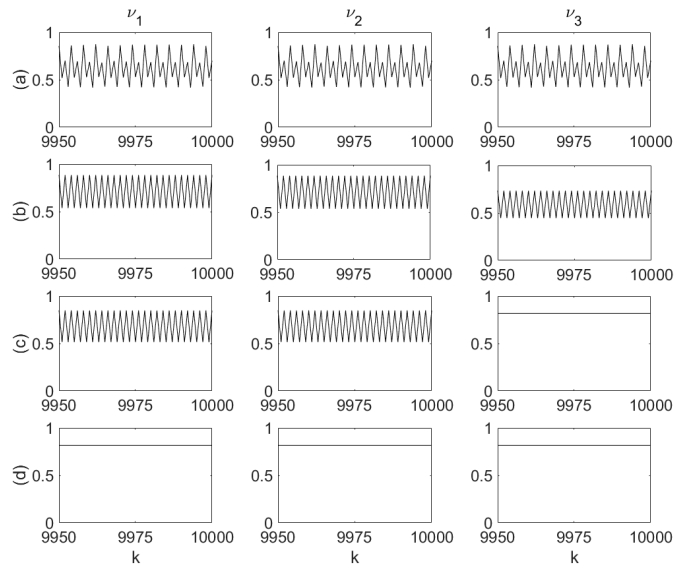 Figure 6. Firing rate sequences corresponding to circuit modification and segregation cases depicted in Figure 5.
Figure 6. Firing rate sequences corresponding to circuit modification and segregation cases depicted in Figure 5.It can be seen in Figure 6 that the fully connected circuit (a) fires synchronously in a largely oscillatory multiplexed mode, whereas the transition to partial connectivity in the modified circuit (b) produces a change of the neuronal firing rate modes, which, while being similarly oscillatory, are unequal in amplitude. The circuit segregation into a two-neuron circuit and one individual neuron (c) results in synchronous oscillation and constant (fixed point) firing modes, respectively, while the segregation into three isolated neurons (d) results in each producing a constant firing rate. The firing modes are in agreement with those predicted by Figure 4 (the transition from a fully connected two-neuron circuit in case 4 (c) to a fully connected three-neuron circuit in case 5 (a) has resulted in the mode changing from oscillatory to largely oscillatory, multiplexing two oscillatory modes).
We have shown that, given internal neuronal property, the circuit connectivity structure defines the circuit firing mode as well. It is therefore justified to view circuit connectivity not only as means for information representation, but also as a manifestation of the function to be performed. The permanence of synapse elimination, as opposed to synapse silencing, makes it particularly relevant to long-term memory and to life-long functional proficiency. Synapse elimination results in circuit modification. It can segregate a synchronous circuit into smaller synchronous sub-circuits, isolated against mutual asynchronous interference. Conversely, the weakening of synapses, eventually resulting in their elimination, may be caused by asynchronous interference between neurons and synapses, as suggested by the Hebbian paradigm. Circuits of identical neurons, but different sizes, fire in different firing rate modes. While we have focused on the firing rate and plasticity dynamics corresponding to maturity, the essential persistence of the map and the corresponding global attractor code of firing rate through different developmental stages suggest the life-long relevance of synapse elimination to circuit formation, modification and function.
This study was supported by the Technion's Roy Matas/Winnipeg Chair in Biomedical Engineering.
The author (Y. Baram) declares the existence of no conflicting interest.
| [1] |
Alberti G (1993) Rank one property for derivatives of functions with bounded variation. Proc Roy Soc Edinburgh Sect A 123: 239-274. doi: 10.1017/S030821050002566X

|
| [2] | Alibert JJ, Bouchitté G (1997) Non-uniform integrability and generalized Young measures. J Convex Anal 4: 129-147. |
| [3] |
Ambrosio L, Coscia A, Dal Maso G (1997) Fine properties of functions with bounded deformation. Arch Ration Mech Anal 139: 201-238. doi: 10.1007/s002050050051
|
| [4] |
Ambrosio L, Dal Maso G (1992) On the relaxation in BV(Ω;Rm) of quasi-convex integrals. J Funct Anal 109: 76-97. doi: 10.1016/0022-1236(92)90012-8
|
| [5] | Ambrosio L, Fusco N, Pallara D (2000) Functions of Bounded Variation and Free-Discontinuity Problems, Oxford University Press. |
| [6] |
Amrouche C, Ciarlet PG, Gratie L, et al. (2006) On Saint Venant's compatibility conditions and Poincaré's lemma. C R Math Acad Sci Paris 342: 887-891. doi: 10.1016/j.crma.2006.03.026
|
| [7] |
Arroyo-Rabasa A, De Philippis G, Hirsch J, et al. (2019) Dimensional estimates and rectifiability for measures satisfying linear PDE constraints. Geom Funct Anal 29: 639-658. doi: 10.1007/s00039-019-00497-1
|
| [8] | Arroyo-Rabasa A, De Philippis G, Rindler F (2017) Lower semicontinuity and relaxation of linear-growth integral functionals under PDE constraints. Adv Calc Var arXiv:1701.02230. |
| [9] | Auslender A, Teboulle M (2003) Asymptotic Cones and Functions in Optimization and Variational Inequalities, Springer. |
| [10] |
Babadjian JF (2015) Traces of functions of bounded deformation. Indiana Univ Math J 64: 1271-1290. doi: 10.1512/iumj.2015.64.5601
|
| [11] |
Ball JM, James RD (1987) Fine phase mixtures as minimizers of energy. Arch Ration Mech Anal 100: 13-52. doi: 10.1007/BF00281246
|
| [12] |
Ball JM, Kirchheim B, Kristensen J (2000) Regularity of quasiconvex envelopes. Calc Var Partial Dif 11: 333-359. doi: 10.1007/s005260000041
|
| [13] | Barroso AC, Fonseca I, Toader R (2000) A relaxation theorem in the space of functions of bounded deformation. Ann Sc Norm Super Pisa Cl Sci 29: 19-49. |
| [14] |
Bellettini G, Coscia A, Dal Maso G (1998) Compactness and lower semicontinuity properties in SBD(Ω). Math Z 228: 337-351. doi: 10.1007/PL00004617
|
| [15] |
Boussaid O, Kreisbeck C, Schlömerkemper A (2019) Characterizations of symmetric polyconvexity. Arch Ration Mech Anal 234: 417-451. doi: 10.1007/s00205-019-01395-4
|
| [16] | Breit D, Diening L, Gmeineder F (2017) On the trace operator for functions of bounded Avariation. arXiv:1707.06804. |
| [17] | Caroccia M, Focardi M, Van Goethem N (2019) On the integral representation of variational functionals on bd. ArXiv:1907.11478. |
| [18] |
Chambolle A, Conti S, Iurlano F (2019) Approximation of functions with small jump sets and existence of strong minimizers of Griffith's energy. J Math Pure Appl 128: 119-139. doi: 10.1016/j.matpur.2019.02.001
|
| [19] |
Chiodaroli E, Feireisl E, Kreml O, et al. (2017) A-free rigidity and applications to the compressible Euler system. Ann Mat Pur Appl 196: 1557-1572. doi: 10.1007/s10231-016-0629-9
|
| [20] |
Conti S, Faraco D, Maggi F (2005) A new approach to counterexamples to L1 estimates: Korn's inequality, geometric rigidity, and regularity for gradients of separately convex functions. Arch Ration Mech Anal 175: 287-300. doi: 10.1007/s00205-004-0350-5
|
| [21] |
Conti S, Focardi M, Iurlano F (2017) Integral representation for functionals defined on sbdp in dimension two. Arch Ration Mech Anal 223: 1337-1374. doi: 10.1007/s00205-016-1059-y
|
| [22] |
Conti S, Focardi M, Iurlano F (2018) Which special functions of bounded deformation have bounded variation?. Proc Roy Soc Edinburgh Sect A 148: 33-50. doi: 10.1017/S030821051700004X
|
| [23] | De Giorgi E (1954), Su una teoria generale della misura (r - 1)-dimensionale in uno spazio ad r dimensioni. Ann Mat Pur Appl 36: 191-213. |
| [24] | De Lellis C (2008) A note on Alberti's rank-one theorem, In: Transport Equations and Multi-D Hyperbolic Conservation Laws, Springer, 61-74. |
| [25] | De Philippis G (2008) On the singular part of measures constrained by linear PDEs and applications, In: European Congress of Mathematics, 833-845. |
| [26] | De Philippis G, Marchese A, Rindler F (2017) On a conjecture of Cheeger, In: Measure Theory in Non-Smooth Spaces, De Gruyter, 145-155. |
| [27] |
De Philippis G, Palmieri L, Rindler F (2018) On the two-state problem for general differential operators. Nonlinear Anal 177: 387-396. doi: 10.1016/j.na.2018.03.015
|
| [28] |
De Philippis G, Rindler F (2016) On the structure of A-free measures and applications. Ann Math 184: 1017-1039. doi: 10.4007/annals.2016.184.3.10
|
| [29] |
De Philippis G, Rindler F (2017) Characterization of generalized Young measures generated by symmetric gradients. Arch Ration Mech Anal 224: 1087-1125. doi: 10.1007/s00205-017-1096-1
|
| [30] | De Philippis G, Rindler F (2018) On the structure of measures constrained by linear PDEs, In: Proceedings of the International Congress of Mathematicians (ICM 2018), 2215-2239. |
| [31] | Ebobisse F (2000) On lower semicontinuity of integral functionals in LD(Ω). Ricerche Mat 49: 65-76. |
| [32] |
Ebobisse F (2005) A lower semicontinuity result for some integral functionals in the space SBD. Nonlinear Anal 62: 1333-1351. doi: 10.1016/j.na.2005.04.036
|
| [33] | Federer H (1969) Geometric Measure Theory, Springer. |
| [34] |
Fleming WH, Rishel R (1960) An integral formula for total gradient variation. Arch Math 11: 218-222. doi: 10.1007/BF01236935
|
| [35] |
Fonseca I, Müller S (1992) Quasi-convex integrands and lower semicontinuity in L1. SIAM J Math Anal 23: 1081-1098. doi: 10.1137/0523060
|
| [36] |
Fonseca I, Müller S (1993) Relaxation of quasiconvex functionals in BV(Ω,Rp) for integrands f (x, u, ▽u). Arch Ration Mech Anal 123: 1-49. doi: 10.1007/BF00386367
|
| [37] |
Fonseca I, Müller S (1999) A-quasiconvexity, lower semicontinuity, and Young measures. SIAM J Math Anal 30: 1355-1390. doi: 10.1137/S0036141098339885
|
| [38] |
Friedrich M (2017) A Korn-type inequality in SBD for functions with small jump sets. Math Mod Meth AppL S 27: 2461-2484. doi: 10.1142/S021820251750049X
|
| [39] |
Friedrich M (2018) A piecewise Korn inequality in S BD and applications to embedding and density results. SIAM J Math Anal 50: 3842-3918. doi: 10.1137/17M1129982
|
| [40] | Friedrich M, Solombrino F (2019) Functionals defined on piecewise rigid functions: Integral representation and Γ-convergence. ArXiv:1904.06305. |
| [41] | Fuchs M, Seregin G (2000) Variational Methods for Problems from Plasticity Theory and for Generalized Newtonian Fluids, Springer. |
| [42] | Gargiulo G, Zappale E (2008) A lower semicontinuity result in SBD. J Convex Anal 15: 191-200. |
| [43] | Gmeineder F, Raita B (2018) On critical Lp-differentiability of BD-maps. arXiv:1802.10364. |
| [44] | Grafakos L (2014) Classical Fourier Analysis, 3 Eds., Springer. |
| [45] |
Kinderlehrer D, Pedregal P (1991) Characterizations of Young measures generated by gradients. Arch Ration Mech Anal 115: 329-365. doi: 10.1007/BF00375279
|
| [46] |
Kinderlehrer D, Pedregal P (1994) Gradient Young measures generated by sequences in Sobolev spaces. J Geom Anal 4: 59-90. doi: 10.1007/BF02921593
|
| [47] |
Kirchheim B, Kristensen J (2011) Automatic convexity of rank-1 convex functions. C R Math Acad Sci Paris 349: 407-409. doi: 10.1016/j.crma.2011.03.013
|
| [48] |
Kirchheim B, Kristensen J (2016) On rank-one convex functions that are homogeneous of degree one. Arch Ration Mech Anal 221: 527-558. doi: 10.1007/s00205-016-0967-1
|
| [49] | Kohn RV (1979) New estimates for deformations in terms of their strains. PhD thesis of Princeton University. |
| [50] |
Kohn RV (1982) New integral estimates for deformations in terms of their nonlinear strains. Arch Ration Mech Anal 78: 131-172. doi: 10.1007/BF00250837
|
| [51] | Kosiba K, Rindler F (2019) On the relaxation of integral functionals depending on the symmetrized gradient. ArXiv:1903.05771. |
| [52] | Kristensen J, Rindler F (2010), Characterization of generalized gradient Young measures generated by sequences in W1,1 and BV. Arch Ration Mech Anal 197: 539-598. |
| [53] | Maggi F (2012) Sets of Finite Perimeter and Geometric Variational Problems, Cambridge: Cambridge University Press. |
| [54] |
Massaccesi A, Vittone D (2019) An elementary proof of the rank-one theorem for BV functions. J Eur Math Soc 21: 3255-3258. doi: 10.4171/JEMS/903
|
| [55] | Matthies H, Strang G, Christiansen E (1979) The saddle point of a differential program, In: Energy Methods in Finite Element Analysis, Wiley, 309-318. |
| [56] | Mattila P (1995) Geometry of Sets and Measures in Euclidean Spaces, Cambridge: Cambridge University Press. |
| [57] |
Müller S (1992) On quasiconvex functions which are homogeneous of degree 1. Indiana U Math J 41: 295-301. doi: 10.1512/iumj.1992.41.41017
|
| [58] | Müller S (1999) Variational models for microstructure and phase transitions, In: Calculus of Variations and Geometric Evolution Problems, Springer, 85-210. |
| [59] | Murat F (1978) Compacité par compensation, Ann Sc Norm Super Pisa Cl Sci 5: 489-507. |
| [60] | Murat F (1979) Compacité par compensation. II, In: Proceedings of the International Meeting on Recent Methods in Nonlinear Analysis (Rome, 1978), Pitagora Editrice Bologna, 245-256. |
| [61] |
Ornstein D (1962) A non-inequality for differential operators in the L1 norm. Arch Ration Mech Anal 11: 40-49. doi: 10.1007/BF00253928
|
| [62] |
Preiss D (1987) Geometry of measures in Rn: distribution, rectifiability, and densities. Ann Math 125: 537-643. doi: 10.2307/1971410
|
| [63] | Rindler F (2011) Lower Semicontinuity and Young Measures for Integral Functionals with Linear Growth, PhD thesis of University of Oxford. |
| [64] |
Rindler F (2011) Lower semicontinuity for integral functionals in the space of functions of bounded deformation via rigidity and Young measures. Arch Ration Mech Anal 202: 63-113. doi: 10.1007/s00205-011-0408-0
|
| [65] | Rindler F (2012) Lower semicontinuity and Young measures in BV without Alberti's Rank-One Theorem. Adv Calc Var 5: 127-159. |
| [66] |
Rindler F (2014) A local proof for the characterization of Young measures generated by sequences in BV. J Funct Anal 266: 6335-6371. doi: 10.1016/j.jfa.2014.03.010
|
| [67] | Rindler F (2018) Calculus of Variations, Springer. |
| [68] | Spector D, Van Schaftingen J (2018) Optimal embeddings into lorentz spaces for some vector differential operators via gagliardo's lemma. ArXiv:1811.02691. |
| [69] | Stein EM (1993) Harmonic Analysis, Princeton University Press. |
| [70] | Suquet PM (1978) Existence et régularité des solutions des équations de la plasticité. C R Acad Sci Paris Sér A 286: 1201-1204. |
| [71] |
Suquet PM (1979) Un espace fonctionnel pour les équations de la plasticité. Ann Fac Sci Toulouse Math 1: 77-87. doi: 10.5802/afst.531
|
| [72] | Tartar L (1979) Compensated compactness and applications to partial differential equations, In: Nonlinear Analysis and Mechanics: Heriot-Watt Symposium, Vol. IV, Pitman, 136-212. |
| [73] | Tartar L (1983) The compensated compactness method applied to systems of conservation laws, In: Systems of Nonlinear Partial Differential Equations, Reidel, 263-285. |
| [74] | Temam R (1983) Problèmes Mathématiques en Plasticité, Montrouge: Gauthier-Villars. |
| [75] |
Temam R, Strang G (1980) Functions of bounded deformation. Arch Ration Mech Anal 75: 7-21. doi: 10.1007/BF00284617
|
| 1. | Yoram Baram, Circuit Polarity Effect of Cortical Connectivity, Activity, and Memory, 2018, 30, 0899-7667, 3037, 10.1162/neco_a_01128 | |
| 2. | Yoram Baram, Probabilistically segregated neural circuits and subcritical linguistics, 2020, 14, 1871-4080, 837, 10.1007/s11571-020-09602-9 | |
| 3. | Yoram Baram, Primal-size neural circuits in meta-periodic interaction, 2021, 15, 1871-4080, 359, 10.1007/s11571-020-09613-6 |

Guido De Philippis, Filip Rindler. Fine properties of functions of bounded deformation-an approach via linear PDEs[J]. Mathematics in Engineering, 2020, 2(3): 386-422. doi: 10.3934/mine.2020018

 DownLoad:
DownLoad: 