Citation: Jasmine S. Ritschard, Lea Amato, Yadhu Kumar, Britta Müller, Leo Meile, Markus Schuppler. The role of the surface smear microbiome in the development of defective smear on surface-ripened red-smear cheese[J]. AIMS Microbiology, 2018, 4(4): 622-641. doi: 10.3934/microbiol.2018.4.622

| [1] | Fox PF, McSweeney PLH (2004) Cheese: An Overview, In: Patrick F. Fox, McSweeney PLH, Cogan TM, et al., Editors, Cheese: chemistry, physics and microbiology, Amsterdam: Elsevier, 1–18. |
| [2] | Bockelmann W (2002) Smear-ripened cheeses, In: Roginski H, Fuquay JW, Fox PF, Editors, Encyclopaedia of Dairy Sciences, London: Academic Press, 391–401. |
| [3] | Bockelmann W (2010) Secondary cheese starter cultures, In: Law BA, Tamime AY, Editors, Technology of cheesemaking, Oxford: Blackwell Publishing Ltd, 193–230. |
| [4] | Brennan NM, Cogan TM, Loessner M, et al. (2004) Bacterial surface-ripened cheeses, In: Fox PF, Mcsweeney PLH, Cogan TM, et al., Editors, Cheese: Chemistry, Physics and Microbiology, Amsterdam: Elsevier, 199–225. |
| [5] | McSweeney PLH, Ottogalli G, Fox PF, et al. (2004) Diversity of cheese varieties: An overview, In: Cheese: Chemistry, Physics and Microbiology, Amsterdam: Elsevier, 1–23. |
| [6] | McSweeney PLH, Fox PF (2004) Metabolism of residual lactose and of lactate and citrate, In: Fox PF, McSweeney PLH, Cogan TM, et al., Editors, Cheese: Chemistry, Physics and Microbiology, Amsterdam: Elsevier, 361–371. |
| [7] | Reps A (1993) Bacterial Surface-Ripened Cheeses, In: Fox PF, Editor, Cheese: Chemistry, Physics and Microbiology, New York: Springer US, 137–172. |
| [8] |
Bockelmann W, Hoppe-Seyler T (2001) The surface flora of bacterial smear-ripened cheeses from cow's and goat's milk. Int Dairy J 11: 307–314. doi: 10.1016/S0958-6946(01)00060-7

|
| [9] |
Monnet C, Bleicher A, Neuhaus K, et al. (2010) Assessment of the anti-listerial activity of microfloras from the surface of smear-ripened cheeses. Food Microbiol 27: 302–310. doi: 10.1016/j.fm.2009.11.009
|
| [10] |
Roth E, Schwenninger SM, Eugster-Meier E, et al. (2011) Facultative anaerobic halophilic and alkaliphilic bacteria isolated from a natural smear ecosystem inhibit Listeria growth in early ripening stages. Int J Food Microbiol 147: 26–32. doi: 10.1016/j.ijfoodmicro.2011.02.032
|
| [11] |
Mounier J, Goerges S, Gelsomino R, et al. (2006) Sources of the adventitious microflora of a smear ripened cheese. J Appl Microbiol 101: 668–681. doi: 10.1111/j.1365-2672.2006.02922.x
|
| [12] |
Rademaker J, Peinhopf M, Rijnen L, et al. (2005) The surface microflora dynamics of bacterial smear-ripened Tilsit cheese determined by T-RFLP DNA population fingerprint analysis. Int Dairy J 15: 785–794. doi: 10.1016/j.idairyj.2004.08.027
|
| [13] |
Bockelmann W, Willems K, Neve H, et al. (2005) Cultures for the ripening of smear cheeses. Int Dairy J 15: 719–732. doi: 10.1016/j.idairyj.2004.08.022
|
| [14] |
Bokulich NA, Mills DA (2013) Facility-specific "house" microbiome drives microbial landscapes of artisan cheesemaking plants. Appl Environ Microb 79: 5214–5223. doi: 10.1128/AEM.00934-13
|
| [15] |
Didienne R, Defargues C, Callon C, et al. (2012) Characteristics of microbial biofilm on wooden vats ("gerles") in PDO Salers cheese. Int J Food Microbiol 156: 91–101. doi: 10.1016/j.ijfoodmicro.2012.03.007
|
| [16] |
Mariani C, Briandet R, Chamba JF, et al. (2007) Biofilm ecology of wooden shelves used in ripening the French raw milk smear cheese Reblochon de Savoie. J Dairy Sci 90: 1653–1661. doi: 10.3168/jds.2006-190
|
| [17] |
Oulahal N, Adt I, Mariani C, et al. (2009) Examination of wooden shelves used in the ripening of a raw milk smear cheese by FTIR spectroscopy. Food Control 20: 658–663. doi: 10.1016/j.foodcont.2008.10.012
|
| [18] | Ndoye B, Rasolofo EA, Lapointe G, et al. (2011) A review of the molecular approaches to investigate the diversity and activity of cheese microbiota. Dairy Sci Technol 91: 1–30. |
| [19] |
Jany JL, Barbier G (2008) Culture-independent methods for identifying microbial communities in cheese. Food Microbiol 25: 839–848. doi: 10.1016/j.fm.2008.06.003
|
| [20] |
Justé A, Thomma BPHJ, Lievens B (2008) Recent advances in molecular techniques to study microbial communities in food-associated matrices and processes. Food Microbiol 25: 745–761. doi: 10.1016/j.fm.2008.04.009
|
| [21] |
Quigley L, O'Sullivan O, Beresford TP, et al. (2011) Molecular approaches to analysing the microbial composition of raw milk and raw milk cheese. Int J Food Microbiol 150: 81–94. doi: 10.1016/j.ijfoodmicro.2011.08.001
|
| [22] | Giraffa G, Carminati D (2008) Molecular techniques in food fermentation: principles and applications, In: Cocolin L, Ercolini D, Editors, Molecular Techniques in the Microbial Ecology of Fermented Foods, Berlin: Springer, 1–30. |
| [23] | O'Sullivan DJ (2000) Methods for analysis of the intestinal microflora. Curr Issues Intest Microbiol 1: 39–50. |
| [24] |
van Hijum SAFT, Vaughan EE, Vogel RF (2012) Application of state-of-art sequencing technologies to indigenous food fermentations. Curr Opin Biotech 24: 1–9. doi: 10.1016/j.ceb.2012.01.012
|
| [25] |
Amato L, Ritschard JS, Kurtz O, et al. (2012) Microbial composition of defect smear-a problem evolving during foil-prepacked storage of red-smear cheeses. Int Dairy J 27: 77–85. doi: 10.1016/j.idairyj.2012.07.012
|
| [26] |
Amato L, Ritschard JS, Sprecher I, et al. (2015) Effect of packaging materials, environmental factors and rRNA transcriptional activity of surface microflora on red-smear cheese defect development. Int Dairy J 41: 50–57. doi: 10.1016/j.idairyj.2014.08.022
|
| [27] |
Koutsoumanis K, Pavlis A, Nychas GJE, et al. (2010) Probabilistic model for Listeria monocytogenes growth during distribution, retail storage, and domestic storage of pasteurized milk. Appl Environ Microb 76: 2181–2191. doi: 10.1128/AEM.02430-09
|
| [28] |
Laguerre O, Hoang HM, Flick D (2013) Experimental investigation and modelling in the food cold chain: Thermal and quality evolution. Trends Food Sci Tech 29: 87–97. doi: 10.1016/j.tifs.2012.08.001
|
| [29] |
Brosius J, Palmer ML, Kennedy PJ, et al. (1978) Complete nucleotide sequence of a 16S ribosomal RNA gene from Escherichia coli. P Natl Acad Sci 75: 4801–4805. doi: 10.1073/pnas.75.10.4801
|
| [30] | Lane DJ (1991) 16S/23S rRNA sequencing. Nucleic Acid Tech Bact Syst 1991: 115–175. |
| [31] |
Weisburg WG, Barns SM, Pelletier DA, et al. (1991) 16S ribosomal DNA amplification for phylogenetic study. J Bacteriol 173: 697–703. doi: 10.1128/jb.173.2.697-703.1991
|
| [32] | Sander A, Ruess M, Bereswill S, et al. (1998) Comparison of different DNA fingerprinting techniques for molecular typing of Bartonella henselae isolates. J Clin Microbiol 36: 2973–2981. |
| [33] | Reysenbach AL, Pace NR (1995) Reliable amplification of hyperthermophilic archaeal 16S rRNA genes by the polymerase chain reaction, In: Robb F, Editor, Archaea: A laboratory manual, Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press, 101–107. |
| [34] |
Takai K, Horikoshi K (2000) Rapid detection and quantification of members of the archaeal community by quantitative PCR using fluorogenic probes. Appl Environ Microb 66: 5066–5072. doi: 10.1128/AEM.66.11.5066-5072.2000
|
| [35] | Hugenholtz P, Pitulle C, Hershberger KL, et al. (1998) Novel division level bacterial diversity in a Yellowstone hot spring. J Bacteriol 180: 366–376. |
| [36] |
Walters WA, Caporaso JG, Lauber CL, et al. (2011) PrimerProspector: De novo design and taxonomic analysis of barcoded polymerase chain reaction primers. Bioinformatics 27: 1159–1161. doi: 10.1093/bioinformatics/btr087
|
| [37] |
Huber JA, Mark-Welch DB, Morrison HG, et al. (2007) Microbial population structures in the deep marine biosphere. Science 318: 97–100. doi: 10.1126/science.1146689
|
| [38] |
Wang Y, Qian PY (2009) Conservative fragments in bacterial 16S rRNA genes and primer design for 16S ribosomal DNA amplicons in metagenomic studies. PLoS One 4: e7401. doi: 10.1371/journal.pone.0007401
|
| [39] |
Claesson MJ, Wang Q, O'Sullivan O, et al. (2010) Comparison of two next-generation sequencing technologies for resolving highly complex microbiota composition using tandem variable 16S rRNA gene regions. Nucleic Acids Res 38: e200. doi: 10.1093/nar/gkq873
|
| [40] |
Liu WT, Mirzabekov AD, Stahl DA (2001) Optimization of an oligonucleotide microchip for microbial identification studies: a non-equilibrium dissociation approach. Environ Microbiol 3: 619–629. doi: 10.1046/j.1462-2920.2001.00233.x
|
| [41] |
DeLong EF (1992) Archaea in coastal marine environments. P Natl Acad Sci USA 89: 5685–5689. doi: 10.1073/pnas.89.12.5685
|
| [42] |
Baker GC, Smith JJ, Cowan DA (2003) Review and re-analysis of domain-specific 16S primers. J Microbiol Meth 55: 541–555. doi: 10.1016/j.mimet.2003.08.009
|
| [43] |
Edgar RC, Haas BJ, Clemente JC, et al. (2011) UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27: 2194–2200. doi: 10.1093/bioinformatics/btr381
|
| [44] |
Wang Q, Garrity GM, Tiedje JM, et al. (2007) Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl Environ Microb 73: 5261. doi: 10.1128/AEM.00062-07
|
| [45] |
Monnet C, Ulve V, Sarthou AS, et al. (2008) Extraction of RNA from cheese without prior separation of microbial cells. Appl Environ Microb 74: 5724–5730. doi: 10.1128/AEM.00716-08
|
| [46] |
Riley J, Butler R, Ogilvie D, et al. (1990) A novel, rapid method for the isolation of terminal sequences from yeast artificial chromosome (YAC) clones. Nucleic Acids Res 18: 2887–2890. doi: 10.1093/nar/18.10.2887
|
| [47] | O'Sullivan D, Cotter P, Giblin L, et al. (2013) Nucleic acid-based approaches to investigate microbial-related cheese quality defects. Front Microbiol 4: 1–15. |
| [48] | Ercolini D (2013) High-throughput sequencing and metagenomics: Moving forward in the culture-independent analysis of food microbial ecology. Appl Environ Microb 79: 3148–3155. |
| [49] | Alegría Á, Szczesny P, Mayo B, et al. (2012) Biodiversity in oscypek, a traditional polish cheese, determined by culture-dependent and -independent approaches. Appl Environ Microb 78: 1890–1898. |
| [50] | Bokulich NA, Mills DA (2013) Improved selection of internal transcribed spacer-specific primers enables quantitative, ultra-high-throughput profiling of fungal communities. Appl Environ Microb 79: 2519–2526. |
| [51] |
Lusk T, Ottesen A, White J, et al. (2012) Characterization of microflora in Latin-style cheeses by next-generation sequencing technology. BMC Microbiol 12: 254. doi: 10.1186/1471-2180-12-254
|
| [52] |
Masoud W, Takamiya M, Vogensen FK, et al. (2011) Characterization of bacterial populations in Danish raw milk cheeses made with different starter cultures by denaturating gradient gel electrophoresis and pyrosequencing. Int Dairy J 21: 142–148. doi: 10.1016/j.idairyj.2010.10.007
|
| [53] |
Masoud WF, Vogensen FK, Lillevang S, et al. (2012) The fate of indigenous microbiota, starter cultures, Escherichia coli, Listeria innocua and Staphylococcus aureus in Danish raw milk and cheeses determined by pyrosequencing and quantitative real time (qRT)-PCR. Int J Food Microbiol 153: 192–202. doi: 10.1016/j.ijfoodmicro.2011.11.014
|
| [54] |
Quigley L, O'Sullivan O, Beresford TP, et al. (2012) High-throughput sequencing for detection of subpopulations of bacteria not previously associated with artisanal cheeses. Appl Environ Microb 78: 5717–5723. doi: 10.1128/AEM.00918-12
|
| [55] | Bockelmann W, Krusch U, Engel G, et al. (1997) The microflora of Tilsit cheese. Part 1. Variability of the smear flora. Mol Nutr Food Res 41: 208–212. |
| [56] |
Deetae P, Bonnarme P, Spinnler H, et al. (2007) Production of volatile aroma compounds by bacterial strains isolated from different surface-ripened French cheeses. Appl Microbiol Biotech 76: 1161–1171. doi: 10.1007/s00253-007-1095-5
|
| [57] |
Montel MC, Buchin S, Mallet A, et al. (2014) Traditional cheeses: Rich and diverse microbiota with associated benefits. Int J Food Microbiol 177: 136–154. doi: 10.1016/j.ijfoodmicro.2014.02.019
|
| [58] |
Quigley L, O'Sullivan O, Stanton C, et al. (2013) The complex microbiota of raw milk. FEMS Microbiol Rev 37: 664–698. doi: 10.1111/1574-6976.12030
|
| [59] |
Delbès C, Ali-Mandjee L, Montel MC (2007) Monitoring bacterial communities in raw milk and cheese by culture-dependent and-independent 16S rRNA gene-based analyses. Appl Environ Microb 73: 1882–1891. doi: 10.1128/AEM.01716-06
|
| [60] |
Oikonomou G, Machado VS, Santisteban C, et al. (2012) Microbial diversity of bovine mastitic milk as described by pyrosequencing of metagenomic 16s rDNA. PLoS One 7: e47671. doi: 10.1371/journal.pone.0047671
|
| [61] |
Schirmer BCT, Heir E, Møretrø T, et al. (2013) Microbial background flora in small-scale cheese production facilities does not inhibit growth and surface attachment of Listeria monocytogenes. J Dairy Sci 96: 6161–6171. doi: 10.3168/jds.2012-6395
|
| [62] |
Schmidt VSJ, Kaufmann V, Kulozik U, et al. (2012) Microbial biodiversity, quality and shelf life of microfiltered and pasteurized extended shelf life (ESL) milk from Germany, Austria and Switzerland. Int J Food Microbiol 154: 1–9. doi: 10.1016/j.ijfoodmicro.2011.12.002
|
| [63] |
Verdier-Metz I, Gagne G, Bornes S, et al. (2012) Cow teat skin, a potential source of diverse microbial populations for cheese production. Appl Environ Microb 78: 326–333. doi: 10.1128/AEM.06229-11
|
| [64] |
White DC, Sutton SD, Ringelberg DB (1996) The genus Sphingomonas: Physiology and ecology. Curr Opin Biotech 7: 301–306. doi: 10.1016/S0958-1669(96)80034-6
|
| [65] |
Zastempowska E, Lassa H (2012) Genotypic characterization and evaluation of an antibiotic resistance of Trueperella pyogenes (Arcanobacterium pyogenes) isolated from milk of dairy cows with clinical mastitis. Vet Microbiol 161: 153–158. doi: 10.1016/j.vetmic.2012.07.018
|
| [66] |
Petri RM, Mapiye C, Dugan ME, et al. (2014) Subcutaneous adipose fatty acid profiles and related rumen bacterial populations of steers fed red clover or grass hay diets containing flax or sunflower-seed. PLoS One 9: e104167. doi: 10.1371/journal.pone.0104167
|
| [67] |
Pitta DW, Pinchak WE, Dowd SE, et al. (2010) Rumen bacterial diversity dynamics associated with changing from bermudagrass hay to grazed winter wheat diets. Microbial Ecol 59: 511–522. doi: 10.1007/s00248-009-9609-6
|
| [68] | Ahn JH, Kim SJ, Weon HY, et al. (2014) Fulvimonas yonginensis sp. nov., isolated from greenhouse soil, and emended description of the genus Fulvimonas. Int J Syst Evol Micr 64: 2758–2762. |
| [69] | Baldani JI, Baldani VLD, Seldin L, et al. (1986) Characterization of Herbaspirillum seropedicae gen. nov., sp. nov., a root-associated nitrogen-fixing bacterium. Int J Syst Bacteriol 36: 86–93. |
| [70] | Lee SD (2013) Humibacter antri sp. nov., an actinobacterium isolated from a natural cave, and emended description of the genus Humibacter. Int J Syst Evol Micr 63: 4315–4319. |
| [71] | Vaz-Moreira I, Nobre MF, Ferreira ACS, et al. (2008) Humibacter albus gen. nov., sp. nov., isolated from sewage sludge compost. Int J Syst Evol Micr 58: 1014–1018. |
| [72] |
Chaban B, Ng SY, Jarrell KF (2006) Archaeal habitats-from the extreme to the ordinary. Can J Microbiol 52: 73–116. doi: 10.1139/w05-147
|
| [73] |
Janssen PH, Kirs M (2008) Structure of the archaeal community of the rumen. Appl Environ Microb 74: 3619–3625. doi: 10.1128/AEM.02812-07
|
| [74] |
Jayashree S, Pushpanathan M, Rajendhran J, et al. (2013) Microbial diversity and phylogeny analysis of buttermilk, a fermented milk product, employing 16S rRNA-based pyrosequencing. Food Biotechnol 27: 213–221. doi: 10.1080/08905436.2013.811084
|
| [75] |
Takai K, Sako Y (1999) A molecular view of archaeal diversity in marine and terrestrial hot water environments. FEMS Microbiol Ecol 28: 177–188. doi: 10.1111/j.1574-6941.1999.tb00573.x
|
| [76] |
Babot JD, Hidalgo M, Arganaraz-Martinez E, et al. (2011) Fluorescence in situ hybridization for detection of classical propionibacteria with specific 16S rRNA-targeted probes and its application to enumeration in Gruyère cheese. Int J Food Microbiol 145: 221–228. doi: 10.1016/j.ijfoodmicro.2010.12.024
|
| [77] |
Cenciarini-Borde C, Courtois S, La SB (2009) Nucleic acids as viability markers for bacteria detection using molecular tools. Future Microbiol 4: 45–64. doi: 10.2217/17460913.4.1.45
|
| [78] | Fey A, Eichler S, Flavier S, et al. (2004) Establishment of a Real-Time PCR-based approach for accurate quantification of bacterial RNA targets in water, using Salmonella as a model organism. Appl Environ Microb 70: 3618–3623. |
| [79] |
Piir K, Paier A, Liiv A, et al. (2011) Ribosome degradation in growing bacteria. EMBO Rep 12: 458–462. doi: 10.1038/embor.2011.47
|
| [80] |
Vendeville A, Larivière D, Fourmentin E (2011) An inventory of the bacterial macromolecular components and their spatial organization. FEMS Microbiol Rev 35: 395–414. doi: 10.1111/j.1574-6976.2010.00254.x
|
| [81] | Bora N, Vancanneyt M, Gelsomino R, et al. (2007) Agrococcus casei sp. nov., isolated from the surfaces of smear-ripened cheeses. Int J Syst Evol Micr 57: 92–97. |
| [82] | Li WJ, Chen HH, Kim CJ, et al. (2005) Nesterenkonia sandarakina sp. nov. and Nesterenkonia lutea sp. nov., novel actinobacteria, and emended description of the genus Nesterenkonia. Int J Syst Evol Micr 55: 463–466. |
| [83] | Mounier J, Gelsomino R, Goerges S, et al. (2005) Surface microflora of four smear-ripened cheeses. Appl Environ Microb 71: 6489–6500. |
| [84] | Brennan NM, Brown R, Goodfellow M, et al. (2001) Microbacterium gubbeenense sp. nov., from the surface of a smear-ripened cheese. Int J Syst Evol Micr 51: 1969–1976. |
| [85] |
Mounier J, Monnet C, Jacques N, et al. (2009) Assessment of the microbial diversity at the surface of Livarot cheese using culture-dependent and independent approaches. Int J Food Microbiol 133: 31–37. doi: 10.1016/j.ijfoodmicro.2009.04.020
|
| [86] |
Rea M, Görges S, Gelsomino R, et al. (2007) Stability of the biodiversity of the surface consortia of Gubbeen, a red-smear cheese. J Dairy Sci 90: 2200–2210. doi: 10.3168/jds.2006-377
|
| [87] |
Vacheyrou M, Normand AC, Guyot P, et al. (2011) Cultivable microbial communities in raw cow milk and potential transfers from stables of sixteen French farms. Int J Food Microbiol 146: 253–262. doi: 10.1016/j.ijfoodmicro.2011.02.033
|
| [88] |
Wolfe BE, Button JE, Santarelli M, et al. (2014) Cheese rind communities provide tractable systems for in situ and in vitro studies of microbial diversity. Cell 158: 422–433. doi: 10.1016/j.cell.2014.05.041
|
| [89] |
Brandl H, Fricker-Feer C, Ziegler D, et al. (2014) Distribution and identification of culturable airborne microorganisms in a Swiss milk processing facility. J Dairy Sci 97: 240–246. doi: 10.3168/jds.2013-7028
|
| [90] |
Gill JJ, Sabour PM, Gong J, et al. (2006) Characterization of bacterial populations recovered from the teat canals of lactating dairy and beef cattle by 16S rRNA gene sequence analysis. FEMS Microbiol Ecol 56: 471–481. doi: 10.1111/j.1574-6941.2006.00091.x
|
| [91] |
Lawson PA, Falsen E, Åkervall E, et al. (1997) Characterization of some Actinomyces-like isolates from human clinical specimens: Reclassification of Actinomyces suis (soltys and spratling) as Actinobaculum suis comb. nov. and description of Actinobaculum schaalii sp. nov. Int J Syst Bacteriol 47: 899–903. doi: 10.1099/00207713-47-3-899
|
| [92] | Lin YC, Uemori K, de Briel DA, et al. (2004) Zimmermannella helvola gen. nov., sp. nov., Zimmermannella alba sp. nov., Zimmermannella bifida sp. nov., Zimmermannella faecalis sp. nov. and Leucobacter albus sp. nov., novel members of the family Microbacteriaceae. Int J Syst Evol Micr 54: 1669–1676. |
| [93] |
Mallet A, Guéguen M, Kauffmann F, et al. (2012) Quantitative and qualitative microbial analysis of raw milk reveals substantial diversity influenced by herd management practices. Int Dairy J 27: 13–21. doi: 10.1016/j.idairyj.2012.07.009
|
| [94] | Maszenan AM, Tay JH, Schumann P, et al. (2005) Quadrisphaera granulorum gen. nov., sp. nov., a Gram-positive polyphosphate-accumulating coccus in tetrads or aggregates isolated from aerobic granules. Int J Syst Evol Micr 55: 1771–1777. |
| [95] | Soddell JA, Stainsby FM, Eales KL, et al. (2006) Millisia brevis gen. nov., sp. nov., an actinomycete isolated from activated sludge foam. Int J Syst Evol Micr 56: 739–744. |
| [96] | Albuquerque L, Tiago I, Rainey FA, et al. (2007) Salirhabdus euzebyi gen. nov., sp. nov., a Gram-positive, halotolerant bacterium isolated from a sea salt evaporation pond. Int J Syst Evol Micr 57: 1566–1571. |
| [97] | Hamada M, Iino T, Tamura T, et al. (2009) Serinibacter salmoneus gen. nov., sp. nov., an actinobacterium isolated from the intestinal tract of a fish, and emended descriptions of the families Beutenbergiaceae and Bogoriellaceae. Int J Syst Evol Micr 59: 2809–2814. |
| [98] | Han SK, Nedashkovskaya OI, Mikhailov VV, et al. (2003) Salinibacterium amurskyense gen. nov., sp. nov., a novel genus of the family Microbacteriaceae from the marine environment. Int J Syst Evol Micr 53: 2061–2066. |
| [99] | Zhang YQ, Schumann P, Yu LY, et al. (2007) Zhihengliuella halotolerans gen. nov., sp. nov., a novel member of the family Micrococcaceae. Int J Syst Evol Micr 57: 1018–1023. |
| [100] |
Ruaro A, Andrighetto C, Torriani S, et al. (2013) Biodiversity and characterization of indigenous coagulase-negative staphylococci isolated from raw milk and cheese of North Italy. Food Microbiol 34: 106–111. doi: 10.1016/j.fm.2012.11.013
|
| [101] |
Bernardet JF, Segers P, Vanca M, et al. (1996) Cutting a gordian knot: Emended classification and description of the genus Flavobacterium, emended description of the family Flavobacteriaceae, and proposal of Flavobacterium hydatis nom. nov. (Basonym, Cytophaga aquatilis Strohl and Tait 1978). Int J Syst Bacteriol 46: 128–148. doi: 10.1099/00207713-46-1-128
|
| [102] | Kämpfer P, Lodders N, Falsen E (2011) Hydrotalea flava gen. nov., sp. nov., a new member of the phylum Bacteroidetes and allocation of the genera Chitinophaga, Sediminibacterium, Lacibacter, Flavihumibacter, Flavisolibacter, Niabella, Niastella, Segetibacter, Parasegetibacter, Terrimonas, Ferruginibacter, Filimonas and Hydrotalea to the family Chitinophagaceae fam. nov. Int J Syst Evol Micr 61: 518–523. |
| [103] |
Deetae P, Mounier J, Bonnarme P, et al. (2009) Effects of Proteus vulgaris growth on the establishment of a cheese microbial community and on the production of volatile aroma compounds in a model cheese. J Appl Microbiol 107: 1404–1413. doi: 10.1111/j.1365-2672.2009.04315.x
|
| [104] |
Maoz A, Mayr R, Scherer S (2003) Temporal stability and biodiversity of two complex antilisterial cheese-ripening microbial consortia. Appl Environ Microb 69: 4012–4018. doi: 10.1128/AEM.69.7.4012-4018.2003
|
| [105] |
Tornadijo M, García M, Fresno J, et al. (2001) Study of Enterobacteriaceae during the manufacture and ripening of San Simón cheese. Food Microbiol 18: 499–509. doi: 10.1006/fmic.2001.0423
|
| [106] |
Irlinger F, Yung SA, Sarthou AS, et al. (2012) Ecological and aromatic impact of two Gram-negative bacteria (Psychrobacter celer and Hafnia alvei) inoculated as part of the whole microbial community of an experimental smear soft cheese. Int J Food Microbiol 153: 332–338. doi: 10.1016/j.ijfoodmicro.2011.11.022
|
| [107] |
Schmitz-Esser S, Dzieciol M, Nischler E, et al. (2018) Abundance and potential contribution of Gram-negative cheese rind bacteria from Austrian artisanal hard cheeses. Int J Food Microbiol 266: 95–103. doi: 10.1016/j.ijfoodmicro.2017.11.013
|
| [108] |
Delbès-Paus C, Pochet S, Helinck S, et al. (2012) Impact of Gram-negative bacteria in interaction with a complex microbial consortium on biogenic amine content and sensory characteristics of an uncooked pressed cheese. Food Microbiol 30: 74–82. doi: 10.1016/j.fm.2011.12.008
|
| [109] | Erkorkmaz BA, Kirtel O, Ateş DÖ, et al. (2018) Development of a cost-effective production process for Halomonas levan. Bioprocess Biosyst Eng 2018: 1–13. |
| [110] |
Finore I, Di Donato P, Mastascusa V, et al. (2014) Fermentation technologies for the optimization of marine microbial exopolysaccharide production. Mar Drugs 12: 3005–3024. doi: 10.3390/md12053005
|
 microbiol-04-04-622-s1.pdf microbiol-04-04-622-s1.pdf |
 |
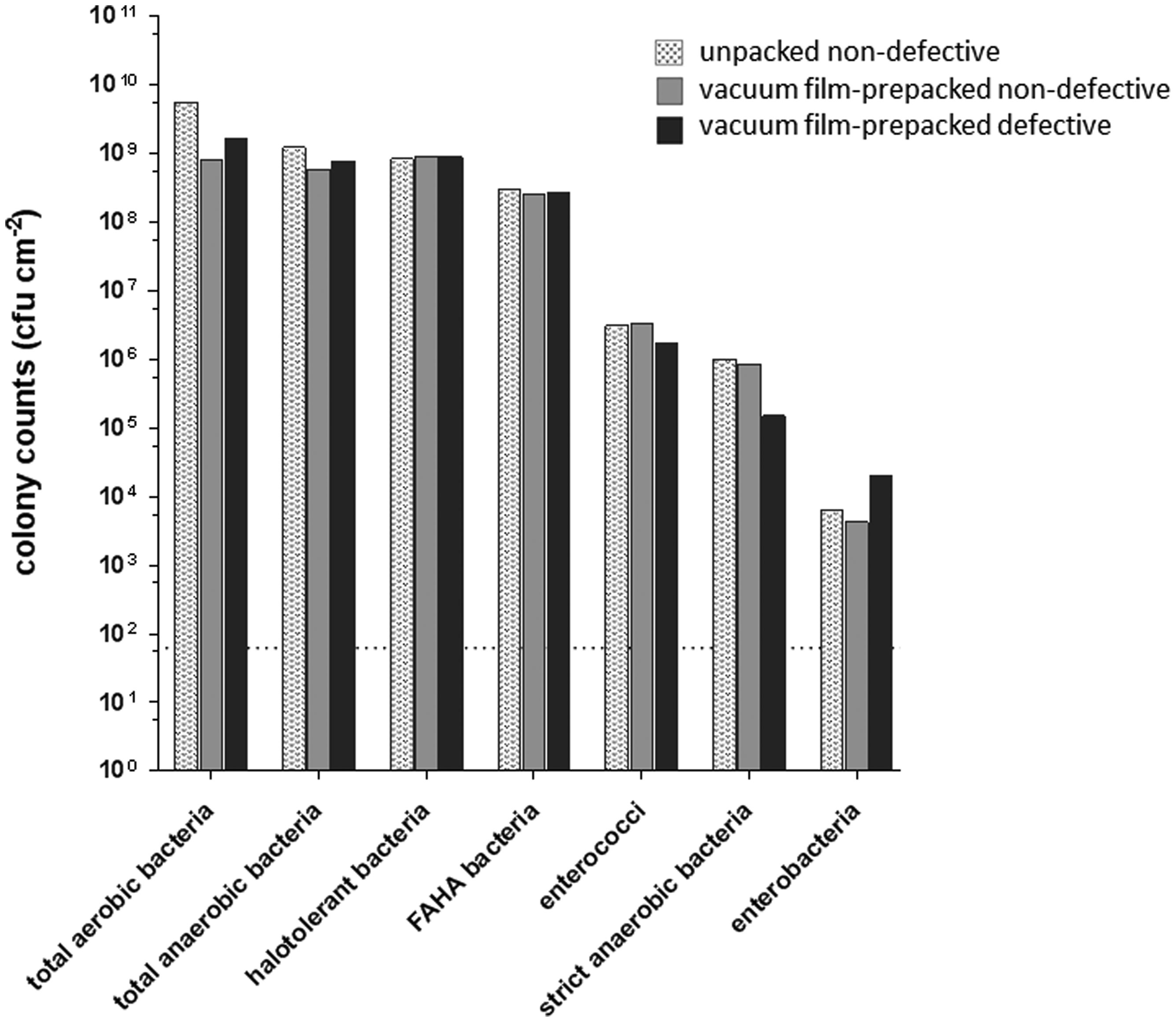
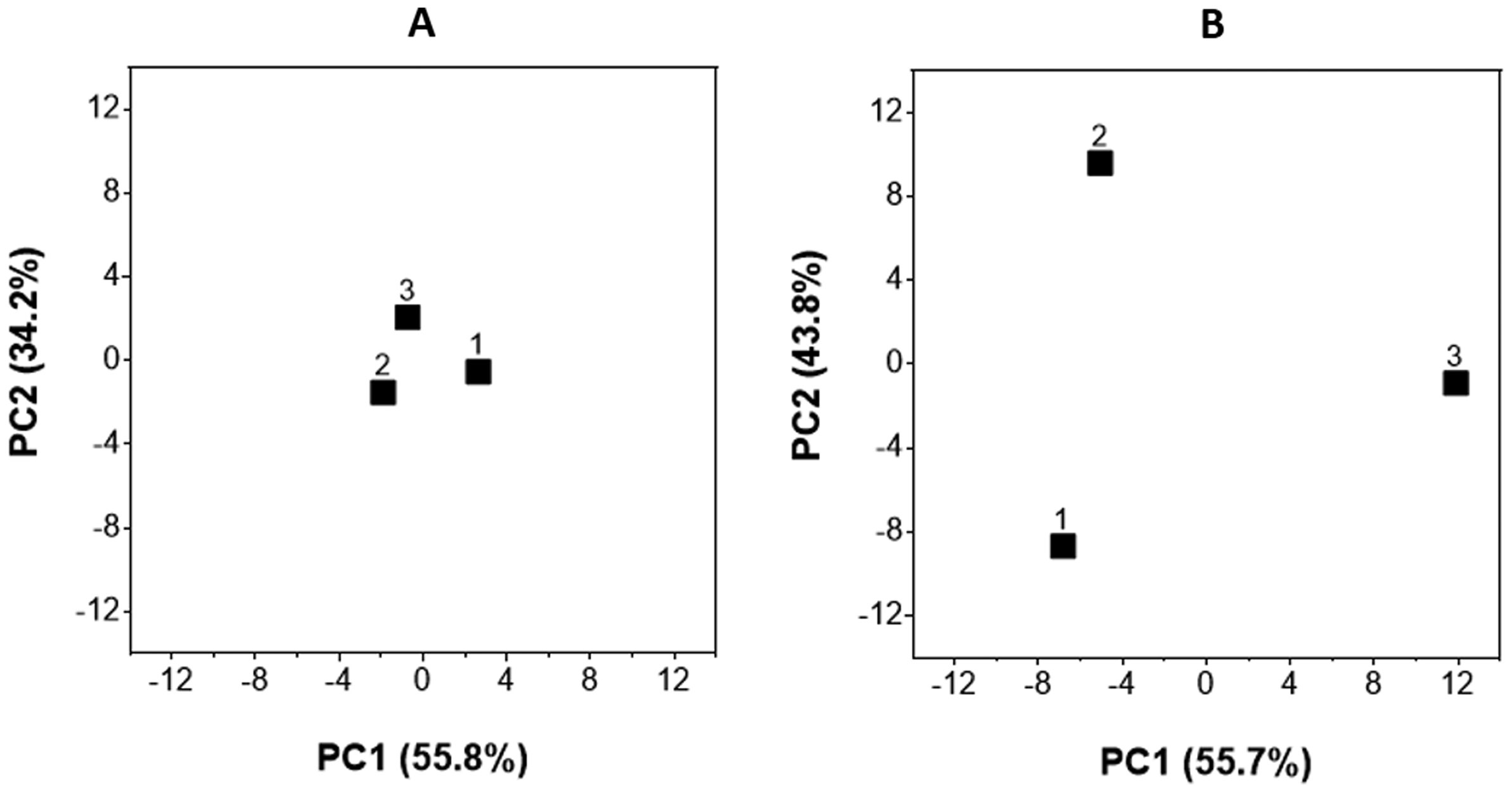
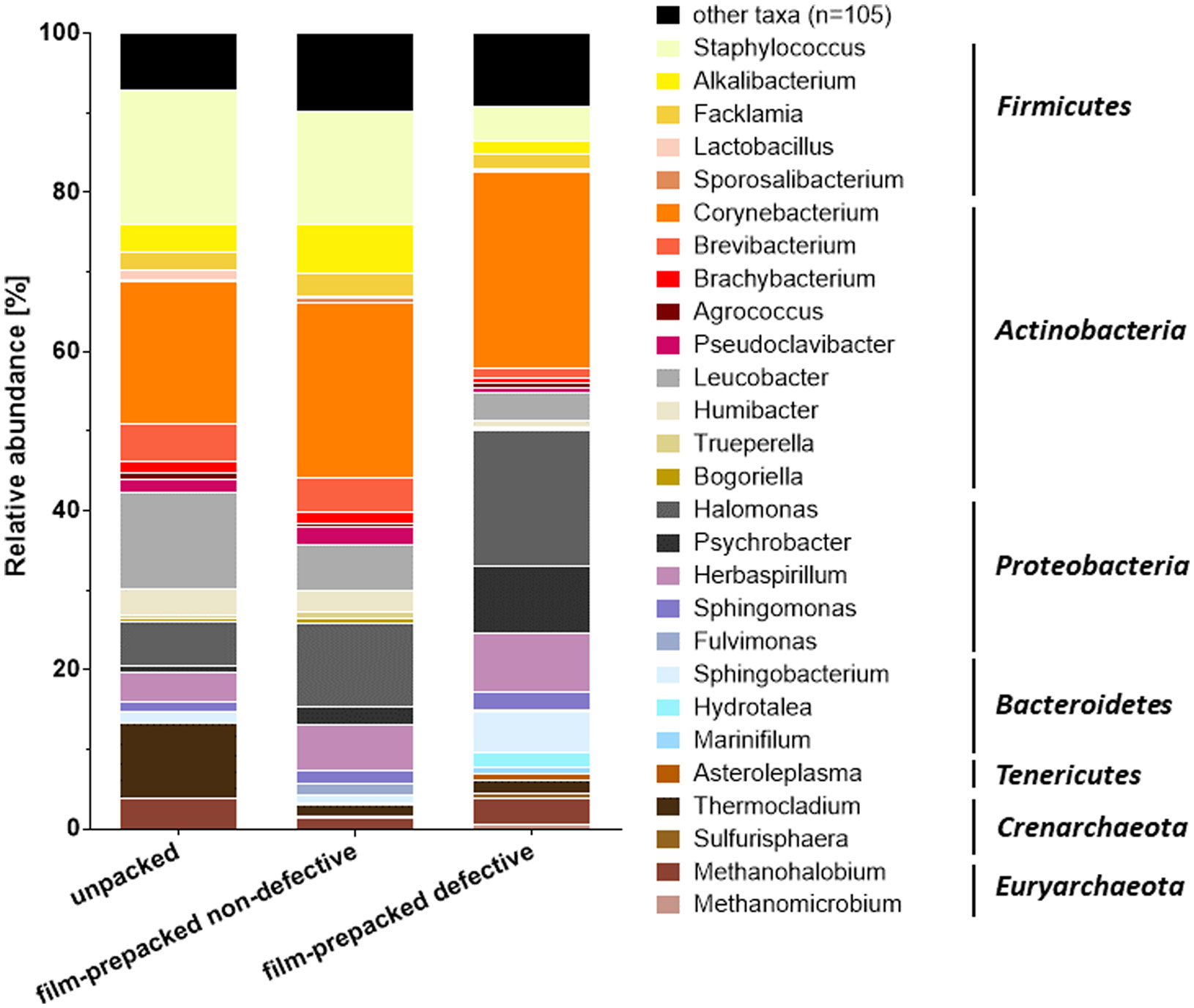
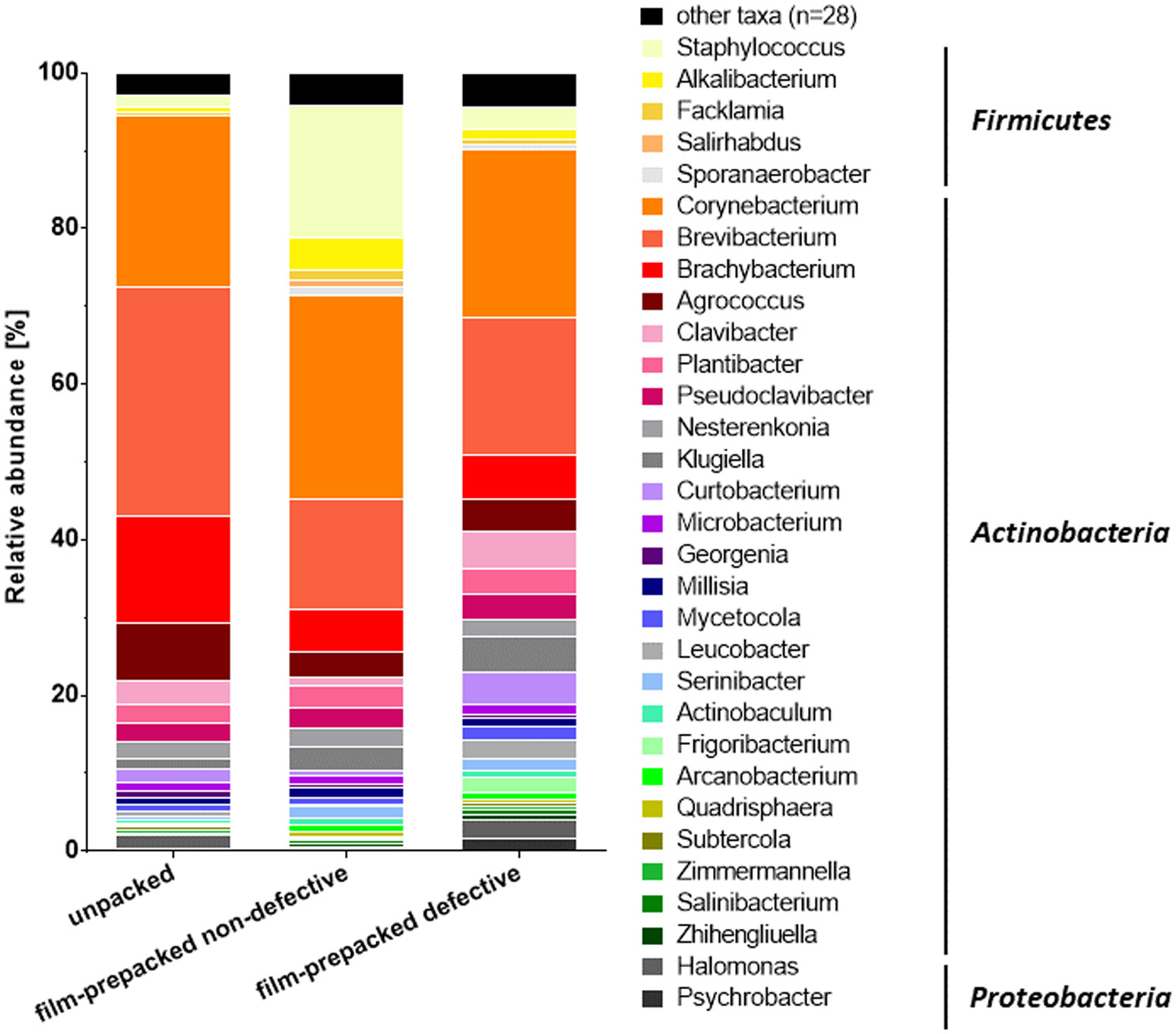
Figures(4) / Tables(1)

Jasmine S. Ritschard, Lea Amato, Yadhu Kumar, Britta Müller, Leo Meile, Markus Schuppler. The role of the surface smear microbiome in the development of defective smear on surface-ripened red-smear cheese[J]. AIMS Microbiology, 2018, 4(4): 622-641. doi: 10.3934/microbiol.2018.4.622







 DownLoad:
DownLoad: