Citation: Stephen T. Abedon. Phage “delay” towards enhancing bacterial escape from biofilms: a more comprehensive way of viewing resistance to bacteriophages[J]. AIMS Microbiology, 2017, 3(2): 186-226. doi: 10.3934/microbiol.2017.2.186

| [1] | Whitman WB, Coleman DC, Wiebe WJ (1998) Prokaryotes: The unseen majority. Proc Natl Acad Sci USA 95: 6578–6583. |
| [2] | Flemming HC, Wingender J (2010) The biofilm matrix. Nat Rev Microbiol 8: 623–633. |
| [3] | Hall MR, McGillicuddy E, Kaplan LJ (2014) Biofilm: basic principles, pathophysiology, and implications for clinicians. Surg Infect (Larchmt ) 15: 1–7. |
| [4] | Hyman P, Abedon ST (2012) Smaller fleas: viruses of microorganisms. Scientifica 2012: 734023. |
| [5] | Wommack KE, Colwell RR (2000) Virioplankton: viruses in aquatic ecosystems. Microbiol Mol Biol Rev 64: 69–114. |
| [6] | Weinbauer MG (2004) Ecology of prokaryotic viruses. FEMS Microbiol Rev 28: 127–181. |
| [7] | Abedon ST (2015) Ecology of anti-biofilm agents I. antibiotics versus bacteriophages. Pharmaceuticals 8: 525–558. |
| [8] | Abedon ST (2015) Ecology of anti-biofilm agents II. bacteriophage exploitation and biocontrol of biofilm bacteria. Pharmaceuticals 8: 559–589. |
| [9] | Briandet R, Lacroix-Gueu P, Renault M, et al. (2008) Fluorescence correlation spectroscopy to study diffusion and reaction of bacteriophages inside biofilms. Appl Environ Microbiol 74: 2135–2143. |
| [10] | Fukuyo M, Sasaki A, Kobayashi I (2012) Success of a suicidal defense strategy against infection in a structured habitat. Sci Rep 2: 238. |
| [11] | Hyman P, Abedon ST (2010) Bacteriophage host range and bacterial resistance. Adv Appl Microbiol 70: 217–248. |
| [12] | Labrie SJ, Samson JE, Moineau S (2010) Bacteriophage resistance mechanisms. Nat Rev Microbiol 8: 317–327. |
| [13] | Dy RL, Richter C, Salmond GP, et al. (2014) Remarkable mechanisms in microbes to resist phage infections. Annu Rev Virol 1: 307–331. |
| [14] | Seed KD (2015) Battling phages: How bacteria defend against viral attack. PLoS Path 11: e1004847. |
| [15] | Parma DH, Snyder M, Sobolevski S, et al. (1992) The rex system of bacteriophage l: tolerance and altruistic cell death. Genes Dev 6: 497–510. |
| [16] | Shub DS (1994) Bacterial viruses. Bacterial altruism? Curr Biol 4: 555–556. |
| [17] | Dy RL, Przybilski R, Semeijn K, et al. (2014) A widespread bacteriophage abortive infection system functions through a Type IV toxin-antitoxin mechanism. Nucl Acids Res 42: 4590–4605. |
| [18] | Iranzo J, Lobkovsky AE, Wolf YI, et al. (2015) Immunity, suicide or both? Ecological determinants for the combined evolution of anti-pathogen defense systems. BMC Evol Biol 15: 43. |
| [19] | Dennehy JJ, Abedon ST, Turner PE (2007) Host density impacts relative fitness of bacteriophage f6 genotypes in structured habitats. Evolution 61: 2516–2527. |
| [20] | Abedon ST, Yin J (2008) Impact of spatial structure on phage population growth, In: Abedon, S.T. Editor, Bacteriophage Ecology, Cambridge, UK: Cambridge University Press, 94–113. |
| [21] | Abedon ST (2011) Bacteriophages and biofilms: ecology, phage therapy, plaques, Hauppauge, New York: Nova Science Publishers. |
| [22] | Abedon ST (2016) Bacteriophage exploitation of bacterial biofilms: phage preference for less mature targets? FEMS Microbiol Lett 363: fnv246. |
| [23] | Abedon ST (2012) Spatial vulnerability: bacterial arrangements, microcolonies, and biofilms as responses to low rather than high phage densities. Viruses 4: 663–687. |
| [24] | Abedon ST (2012) Thinking about microcolonies as phage targets. Bacteriophage 2: 200–204. |
| [25] | Abedon ST (2008) Phage population growth: constraints, games, adaptation, In: Abedon, S.T. Editor, Bacteriophage Ecology, Cambridge, UK: Cambridge University Press, 64–93. |
| [26] | Abedon ST (2009) Impact of phage properties on bacterial survival, In: Adams, H.T. Editor, Contemporary Trends in Bacteriophage Research, Hauppauge, New York: Nova Science Publishers, 217–235. |
| [27] | Payne RJH, Jansen VAA (2001) Understanding bacteriophage therapy as a density-dependent kinetic process. J Theor Biol 208: 37–48. |
| [28] | Abedon ST, Thomas-Abedon C (2010) Phage therapy pharmacology. Curr Pharm Biotechnol 11: 28–47. |
| [29] | Hamilton WD (1964) The genetical evolution of social behaviour. I. J Theor Biol 7: 1–16. |
| [30] | West SA, Gardner A (2010) Altruism, spite, and greenbeards. Science 327: 1341–1344. |
| [31] | Iranzo J, Lobkovsky AE, Wolf YI, et al. (2014) Virus-host arms race at the joint origin of multicellularity and programmed cell death. Cell Cycle 13: 3083–3088. |
| [32] | West SA, Griffin AS, Gardner A, et al. (2006) Social evolution theory for microorganisms. Nat Rev Microbiol 4: 597–607. |
| [33] | Ewald PW (1994) Evolution of Infectious Disease, New York: Oxford University Press. |
| [34] | Michod RE, Nedelcu AM, Roze D (2003) Cooperation and conflict in the evolution of individuality. IV. Conflict mediation and evolvability in Volvox carteri. Bio Systems 69: 95–114. |
| [35] | van Gestel J, Vlamakis H, Kolter R (2015) Division of labor in biofilms: the ecology of cell differentiation. Microbiol Spectr 3: MB-0002-2014. |
| [36] | Andrews JH (1998) Bacteria as modular organisms. Ann Rev Microbiol 52: 105–126. |
| [37] | Kaplan JB (2010) Biofilm dispersal: mechanisms, clinical implications, and potential therapeutic uses. J Dent Res 89: 205–218. |
| [38] | McDougald D, Rice SA, Barraud N, et al. (2012) Should we stay or should we go: mechanisms and ecological consequences for biofilm dispersal. Nat Rev Microbiol 10: 39–50. |
| [39] | Davies DG (2011) Biofilm disperson, In: Flemming, H.C., Wingender, J., Szewzyk, U. Editors, Biofilm Highlights, Berlin: Springer, 1–28. |
| [40] | Barraud N, Kjelleberg S, Rice SA (2015) Dispersal from microbial biofilms. Microbiol Spectr 3: MB-0015-2014. |
| [41] | Petrova OE, Sauer K (2016) Escaping the biofilm in more than one way: desorption, detachment or dispersion. Curr Opin Microbiol 30: 67–78. |
| [42] | Kim SK, Lee JH (2016) Biofilm dispersion in Pseudomonas aeruginosa. J Microbiol 54: 71–85. |
| [43] | Ronce O (2007) How does it feel to be like a rolling stone? Ten questions about dispersal evolution. Ann Rev Ecol Evol Syst 38: 231–253. |
| [44] | Korber DR, Lawrence JR, Lappin-Scott HM, et al. (1995) Growth of microorganisms on surfaces, In: Costerton, J.W., Lappin-Scott, H. Editors, Microbial biofilms, Cambridge, UK: Cambridge University Press, 15–45. |
| [45] | Bjarnsholt T (2013) The role of bacterial biofilms in chronic infections. Apmis 121: 1–51. |
| [46] | Battin TJ, Besemer K, Bengtsson MM, et al. (2016) The ecology and biogeochemistry of stream biofilms. Nat Rev Microbiol 14: 251–263. |
| [47] | Costerton JW, Lappin-Scott H (1995) Introduction to microbial biofilms, In: Costerton, J.W., Lappin-Scott, H. Editors, Microbial biofilms, Cambridge, UK: Cambridge University Press, 1–11. |
| [48] | Costerton JW, Marrie TJ, Cheng KJ (1985) Phenomena of bacterial adhesion, In: Savage, D.C., Fletcher, M. Editors, Bacterial Adhesion: Mechanisms and Physiological Significance, New York: Plenum Press, 3–43. |
| [49] | Somerville DA, Noble WC (1973) Microcolony size of microbes on human skin. J Med Microbiol 6: 323–328. |
| [50] | Mitarai N, Brown S, Sneppen K (2016) Population dynamics of phage and bacteria in spatially structured habitats using phage l and Escherichia coli. J Bacteriol 198: 1783–1793. |
| [51] | Kreft JU (2004) Biofilms promote altruism. Microbiology 150: 2751–2760. |
| [52] | Nadell CD, Bassler BL (2011) A fitness trade-off between local competition and dispersal in Vibrio cholerae biofilms. Proc Natl Acad Sci USA 108: 14181–14185. |
| [53] | Bryers JD (1988) Modeling biofilm accumulation, In: Brazin, M.J., Prosser, J.I. Editors, Physiological Models in Microbiology, Eds., FL: CRC Press, 109–144. |
| [54] | Garny K, Horn H, Neu TR (2008) Interaction between biofilm development, structure and detachment in rotating annular reactors. Bioprocess Biosyst Eng 31: 619–629. |
| [55] | Brading MG, Jass J, Lappin-Scott HM (1995) Dynamics of bacterial biofilm formation, In: Costerton, J.W., Lappin-Scott, H. Editors, Microbial biofilms, Cambridge, UK: Cambridge University Press, 46–63. |
| [56] | Vanysacker L, Boerjan B, Declerck P, et al. (2014) Biofouling ecology as a means to better understand membrane biofouling. Appl Microbiol Biotechnol 98: 8047–8072. |
| [57] | Bester E, Wolfaardt G, Joubert L, et al. (2005) Planktonic-cell yield of a pseudomonad biofilm. Appl Environ Microbiol 71: 7792–7798. |
| [58] | Knowles B, Silveira CB, Bailey BA, et al. (2016) Lytic to temperate switching of viral communities. Nature 531: 466–470. |
| [59] | Gonzalez S, Fernandez L, Campelo AB, et al. (2017) The behavior of Staphylococcus aureus dual-species biofilms treated with bacteriophage phiIPLA-RODI depends on the accompanying microorganism. Appl Environ Microbiol 83: AEM-02821-16. |
| [60] | Sutherland IW, Hughes KA, Skillman LC, et al. (2004) The interaction of phage and biofilms. FEMS Microbiol Lett 232: 1–6. |
| [61] | Nale JY, Chutia M, Carr P, et al. (2016) "Get in early"; biofilm and wax moth (Galleria mellonella) models reveal new insights into the therapeutic potential of Clostridium difficile bacteriophages. Front Microbiol 7: 1383. |
| [62] | Doolittle MM, Cooney JJ, Caldwell DE (1996) Tracing the interaction of bacteriophage with bacterial biofilms using fluorescent and chromogenic probes. J Indust Microbiol 16: 331–341. |
| [63] | Abedon ST (1992) Lysis of lysis inhibited bacteriophage T4-infected cells. J Bacteriol 174: 8073–8080. |
| [64] | Abedon ST (1989) Selection for bacteriophage latent period length by bacterial density: A theoretical examination. Microb Ecol 18: 79–88. |
| [65] | Wang IN, Dykhuizen DE, Slobodkin LB (1996) The evolution of phage lysis timing. Evol Ecol 10: 545–558. |
| [66] | Hadas H, Einav M, Fishov I, et al. (1997) Bacteriophage T4 development depends on the physiology of its host Escherichia coli. Microbiology 143: 179–185. |
| [67] | Abedon ST (2012) Bacterial "immunity" against bacteriophages. Bacteriophage 2: 50–54. |
| [68] | Koonin EV, Zhang F (2017) Coupling immunity and programmed cell suicide in prokaryotes: Life-or-death choices. BioEssays 39: 1–9. |
| [69] | Hoyland-Kroghsbo NM, Paczkowski J, Mukherjee S, et al. (2017) Quorum sensing controls the Pseudomonas aeruginosa CRISPR-Cas adaptive immune system. Proc Natl Acad Sci USA 114: 131–135. |
| [70] | Goldfarb T, Sberro H, Weinstock E, et al. (2015) BREX is a novel phage resistance system widespread in microbial genomes. EMBO J 34: 169–183. |
| [71] | Abedon ST (2015) Bacteriophage secondary infection. Virol Sin 30: 3–10. |
| [72] | McLean RJ, Corbin BD, Balzer GJ, et al. (2001) Phenotype characterization of genetically defined microorganisms and growth of bacteriophage in biofilms. Meth Enzymol 336: 163–174. |
| [73] | Azeredo J, Sutherland IW (2008) The use of phages for the removal of infectious biofilms. Curr Pharm Biotechnol 9: 261–266. |
| [74] | Sillankorva S, Azeredo J (2014) The use of bacteriophages and bacteriophage-derived enzymes for clinically relevant biofilm control, In: Borysowski, J., Miedzybrodzki, R., Górski, A. Editors, Phage Therapy: Current Research and Applications, Norfolk, UK: Caister Academic Press, 305–325. |
| [75] | Kutter E, Kellenberger E, Carlson K, et al. (1994) Effects of bacterial growth conditions and physiology on T4 infection, In: Karam, J.D., Kutter, E., Carlson, K., Guttman, B. Editors, The Molecular Biology of Bacteriophage T4, Washington, DC: ASM Press, 406–418. |
| [76] | Miller RV, Day M (2008) Contribution of lysogeny, pseudolysogeny, and starvation to phage ecology, In: Abedon, S.T. Editor, Bacteriophage Ecology, Cambridge, UK: Cambridge University Press, 114–143. |
| [77] | Pearl S, Gabay C, Kishony R, et al. (2008) Nongenetic individuality in the host-phage interaction. PLoS Biol 6: e120. |
| [78] | Abedon ST (2009) Disambiguating bacteriophage pseudolysogeny: an historical analysis of lysogeny, pseudolysogeny, and the phage carrier state, In: Adams, H.T. Editor, Contemporary Trends in Bacteriophage Research, Hauppauge, New York: Nova Science Publishers, 285–307. |
| [79] | Los M, Wegrzyn G (2012) Pseudolysogeny. Adv Virus Res 82: 339–349. |
| [80] | Golec P, Karczewska-Golec J, Los M, et al. (2014) Bacteriophage T4 can produce progeny virions in extremely slowly growing Escherichia coli host: comparison of a mathematical model with the experimental data. FEMS Microbiol Lett 351: 156–161. |
| [81] | Harper DR, Parracho HMR, Walker J, et al. (2014) Bacteriophages and biofilms. Antibiotics 3: 270–284. |
| [82] | Bryan D, el-Shibiny A, Hobbs Z, et al. (2016) Bacteriophage T4 infection of stationary phase E. coli: life after log from a phage perspective. Front Microbiol 7: 1391. |
| [83] | Yin J (1991) A quantifiable phenotype of viral propagation. Biochem Biophys Res Com 174: 1009–1014. |
| [84] | Filippini M, Buesing N, Bettarel Y, et al. (2006) Infection paradox: high abundance but low impact of freshwater benthic viruses. Appl Environ Microbiol 72: 4893–4898. |
| [85] | Wilkinson JF (1958) The extracellualr polysaccharides of bacteria. Bacteriol Rev 22: 46–73. |
| [86] | van Benthum WAJ, van Loosdrecht MCM, Tijhuis L, et al. (1995) Solids retention time in heterotrophic and nitrifying biofilms in a biofilm airlift suspension reactor. Water Sci Technol 32: 35–43. |
| [87] | Reichert P, Wanner O (1997) Movement of solids in biofilms-significance of liquid phase transport. Water Sci Technol 36: 321–328. |
| [88] | Davey ME, O'Toole GA (2000) Microbial biofilms: from ecology to molecular genetics. Microbiol Mol Biol Rev 64: 847–867. |
| [89] | Hanlon GW, Denyer SP, Olliff CJ, et al. (2001) Reduction in exopolysaccharide viscosity as an aid to bacteriophage penetration through Pseudomonas aeruginosa biofilms. Appl Environ Microbiol 67: 2746–2753. |
| [90] | Resch A, Fehrenbacher B, Eisele K, et al. (2005) Phage release from biofilm and planktonic Staphylococcus aureus cells. FEMS Microbiol Lett 252: 89–96. |
| [91] | Forde A, Fitzgerald GF (2003) Molecular organization of exopolysaccharide (EPS) encoding genes on the lactococcal bacteriophage adsorption blocking plasmid, pCI658. Plasmid 49: 130–142. |
| [92] | Flood JA, Ashbolt NJ (2000) Virus-sized particles can be entrapped and concentrated one hundred fold within wetland biofilms. Adv Environ Res 3: 403–411. |
| [93] | Storey MV, Ashbolt NJ (2001) Persistence of two model enteric viruses (B40-8 and MS-2 bacteriophages) in water distribution pipe biofilms. Water Sci Technol 43: 133–138. |
| [94] | Lacroix-Gueu P, Briandet R, Lévêque-Fort S, et al. (2005) In situ measurements of viral particles diffusion inside mucoid biofilms. C R Biol 328: 1065–1072. |
| [95] | Brüssow H (2013) Bacteriophage-host interaction: from splendid isolation into a messy reality. Curr Opin Microbiol 16: 500–506. |
| [96] | Fan X, Li W, Zheng F, et al. (2013) Bacteriophage inspired antibiotics discovery against infection involved biofilm. Crit Rev Eukaryot Gene Expr 23: 317–326. |
| [97] | Parasion S, Kwiatek M, Gryko R, et al. (2014) Bacteriophages as an alternative strategy for fighting biofilm development. Pol J Microbiol 63: 137–145. |
| [98] | Chan BK, Abedon ST (2015) Bacteriophages and their enzymes in biofilm control. Curr Pharm Des 21: 85–99. |
| [99] | Gutierrez D, Rodriguez-Rubio L, Martinez B, et al. (2016) Bacteriophages as weapons against bacterial biofilms in the food industry. Front Microbiol 7: 825. |
| [100] | Khalifa L, Shlezinger M, Beyth S, et al. (2016) Phage therapy against Enterococcus faecalis in dental root canals. J Oral Microbiol 8: 32157. |
| [101] | Motlagh AM, Bhattacharjee AS, Goel R (2016) Biofilm control with natural and genetically-modified phages. World J Microbiol Biotechnol 32: 67. |
| [102] | Abedon ST (2014) Phage therapy: eco-physiological pharmacology. Scientifica 2014: 581639. |
| [103] | Arber W, Linn S (1969) DNA modification and restriction. Annu Rev Biochem 38: 467–500. |
| [104] | Korona R, Levin BR (1993) Phage-mediated selection and the evolution and maintenance of restriction-modification. Evolution 47: 556–575. |
| [105] | Chao L, Levin BR, Stewart FM (1977) A complex community in a simple habitat: an experimental study with bacteria and phage. Ecology 58: 369–378. |
| [106] | Bohannan BJM, Travisano M, Lenski RE (1999) Epistatic interactions can lower the cost of resistance to multiple consumers. Evolution 53: 292–295. |
| [107] | Hill C (1993) Bacteriophage and bacteriophage resistance in lactic acid bacteria. FEMS Microbiol Rev 12: 87–108. |
| [108] | Nissen SB, Magidson T, Gross K, et al. (2016) Publication bias and the canonization of false facts. eLife 5: e21451. |
| [109] | Costerton JW, Cheng JJ, Geesey GG, et al. (1987) Bacterial biofilms in nature and disease. Ann Rev Microbiol 41: 435–464. |
| [110] | Oh YJ, Jo W, Yang Y, et al. (2007) Influence of culture conditions on Escherichia coli O157:H7 biofilm formation by atomic force microscopy. Ultramicroscopy 107: 869–874. |
| [111] | Cheng KJ, Costerton JW (1980) The formation of microcolonies by rumen bacteria. Can J Microbiol 26: 1104–1113. |
| [112] | Lacqua A, Wanner O, Colangelo T, et al. (2006) Emergence of biofilm-forming subpopulations upon exposure of Escherichia coli to environmental bacteriophages. Appl Environ Microbiol 72: 956–959. |
| [113] | Shen Y, Mitchell MS, Donovan DM, et al. (2012) Phage-based enzybiotics, In: Hyman, P., Abedon, S.T. Editors, Bacteriophages in Health and Disease, Wallingford, UK: CABI Press, 217–239. |
| [114] | Yan J, Mao J, Xie J (2014) Bacteriophage polysaccharide depolymerases and biomedical applications. BioDrugs 28: 265–274. |
| [115] | Gutiérrez D, Briers Y, Rodríguez-Rubio L, et al. (2015) Role of the pre-neck appendage protein (Dpo7) from phage vB_SepiS-phiIPLA7 as an anti-biofilm agent in staphylococcal species. Front Microbiol 6: 1315. |
| [116] | Pires DP, Oliveira H, Melo LD, et al. (2016) Bacteriophage-encoded depolymerases: their diversity and biotechnological applications. Appl Microbiol Biotechnol 100: 2141–2151. |
| [117] | Westra ER, van HS, Oyesiku-Blakemore S, et al. (2015) Parasite exposure drives selective evolution of constitutive versus inducible defense. Curr Biol 25: 1043–1049. |
| [118] | Berngruber TW, Lion S, Gandon S (2013) Evolution of suicide as a defence strategy against pathogens in a spatially structured environment. Ecol Lett 16: 446–453. |
| [119] | Refardt D, Kummerli R (2013) Defying bacteriophages: contrasting altruistic with individual-based resistance mechanisms in Escherichia coli. Commun Integr Biol 6: e25159. |
| [120] | Heilmann S, Sneppen K, Krishna S (2012) Coexistence of phage and bacteria on the boundary of self-organized refuges. Proc Natl Acad Sci USA 109: 12828–12833. |
| [121] | Abedon ST, Kuhl SJ, Blasdel BG, et al. (2011) Phage treatment of human infections. Bacteriophage 1: 66–85. |
| [122] | Borysowski J, Miedzybrodzki R, Górski A (2014) Phage Therapy: Current Research and Applications, Norfolk, UK: Caister Academic Press. |
| [123] | Scali C, Kunimoto B (2013) An update on chronic wounds and the role of biofilms. J Cutan Med Surg 17: 371–376. |
| [124] | Cooper RA, Bjarnsholt T, Alhede M (2014) Biofilms in wounds: a review of present knowledge. J Wound Care 23: 570–580. |
| [125] | Macia MD, Rojo-Molinero E, Oliver A (2014) Antimicrobial susceptibility testing in biofilm-growing bacteria. Clin Microbiol Infect 20: 981–990. |
| [126] | Abedon ST (2015) Phage therapy of pulmonary infections. Bacteriophage 5: e1020260. |
| [127] | Fish R, Kutter E, Wheat G, et al. (2016) Bacteriophage treatment of intransigent diabetic toe ulcers: a case series. J Wound Care 25: S27–S33. |
| [128] | Abedon ST (2017) Bacteriophage clinical use as antibactertial "drugs": utility precident. Microbiol Spectr. |
| [129] | Stewart PS, Franklin MJ (2008) Physiological heterogeneity in biofilms. Nat Rev Microbiol 6: 199–210. |
| [130] | Davis KM, Mohammadi S, Isberg RR (2015) Community behavior and spatial regulation within a bacterial microcolony in deep tissue sites serves to protect against host attack. Cell Host Microbe 17: 21–31. |
| [131] | Harmsen M, Yang L, Pamp SJ, et al. (2010) An update on Pseudomonas aeruginosa biofilm formation, tolerance, and dispersal. FEMS Immunol Med Microbiol 59: 253–268. |
| [132] | Purevdorj-Gage B, Costerton WJ, Stoodley P (2005) Phenotypic differentiation and seeding dispersal in non-mucoid and mucoid Pseudomonas aeruginosa biofilms. Microbiology 151: 1569–1576. |
| [133] | Hamilton WA (1987) Biofilms: microbial interactions and metabolic activities, In: Fletcher, M., Gray, T.R.G., Jones, J.G. Editors, Ecology of Microbial Communities, Cambridge: Cambridge University Press, 361–385. |
| [134] | Abedon ST, Herschler TD, Stopar D (2001) Bacteriophage latent-period evolution as a response to resource availability. Appl Environ Microbiol 67: 4233–4241. |
| [135] | Levin BR, Bull JJ (2004) Population and evolutionary dynamics of phage therapy. Nat Rev Microbiol 2: 166–173. |
| [136] | Gill JJ (2008) Modeling of bacteriophage therapy, In: Abedon, S.T. Editor, Bacteriophage Ecology, Cambridge, UK: Cambridge University Press, 439–464. |
| [137] | Stent GS (1963) Molecular Biology of Bacterial Viruses, San Francisco, CA: WH Freeman and Co. |
| [138] | Gallet R, Shao Y, Wang IN (2009) High adsorption rate is detrimental to bacteriophage fitness in a biofilm-like environment. BMC Evol Biol 9: 241. |
| [139] | Yin J, McCaskill JS (1992) Replication of viruses in a growing plaque: a reaction-diffusion model. Biophys J 61: 1540–1549. |
| [140] | Abedon ST, Culler RR (2007) Bacteriophage evolution given spatial constraint. J Theor Biol 248: 111–119. |
| [141] | Hewson I, Fuhrman JA (2003) Viriobenthos production and virioplankton sorptive scavenging. Microb Ecol 46: 337–347. |
| [142] | Hoyland-Kroghsbo NM, Maerkedahl RB, Svenningsen SL (2013) A quorum-sensing-induced bacteriophage defense mechanism. MBio 4: e00362. |
| [143] | Boots M, Mealor M (2007) Local interactions select for lower pathogen infectivity. Science 315: 1284–1286. |
| [144] | Abedon ST (2016) Commentary: phage therapy of staphylococcal chronic osteomyelitis in experimental animal model. Front Microbiol 7: 1251. |
| [145] | Kutter E, De Vos D, Gvasalia G, et al. (2010) Phage therapy in clinical practice: treatment of human infections. Curr Pharm Biotechnol 11: 69–86. |
| [146] | Hyman P, Abedon ST (2009) Practical methods for determining phage growth parameters. Meth Mol Biol 501: 175–202. |
| [147] | Mirzaei MK, Nilsson AS (2015) Isolation of phages for phage therapy: a comparison of spot tests and efficiency of plating analyses for determination of host range and efficacy. PLoS One 10: e0118557. |
| [148] | Abedon ST (2014) Bacteriophages as drugs: the pharmacology of phage therapy, In: Borysowski, J., Miedzybrodzki, R., Górski, A. Editors, Phage Therapy: Current Research and Applications, Norfolk, UK: Caister Academic Press, 69–100. |

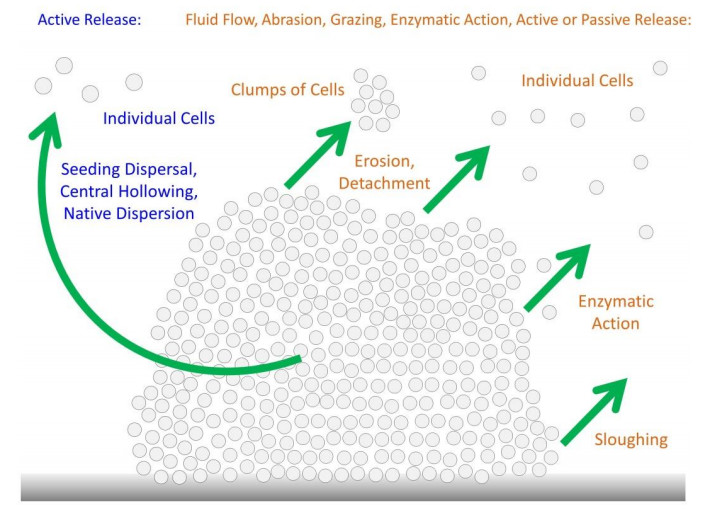
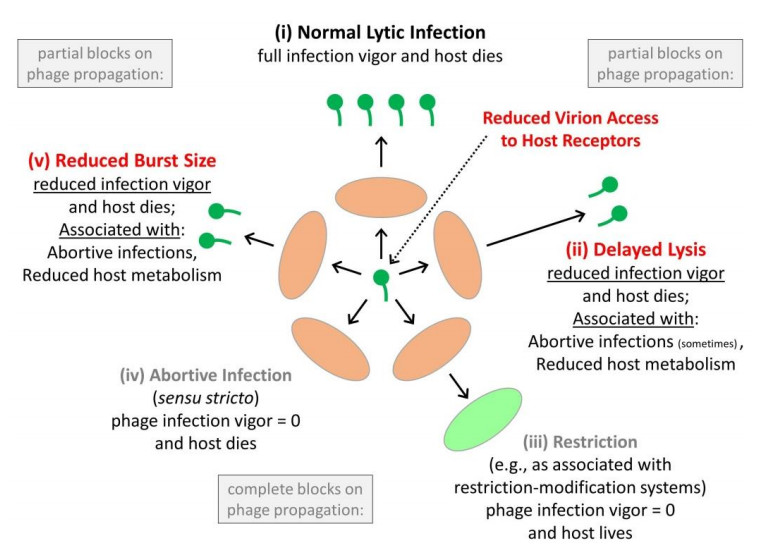
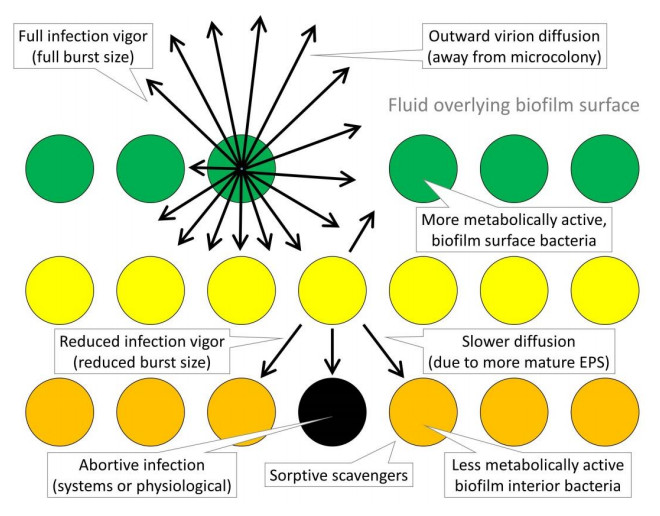
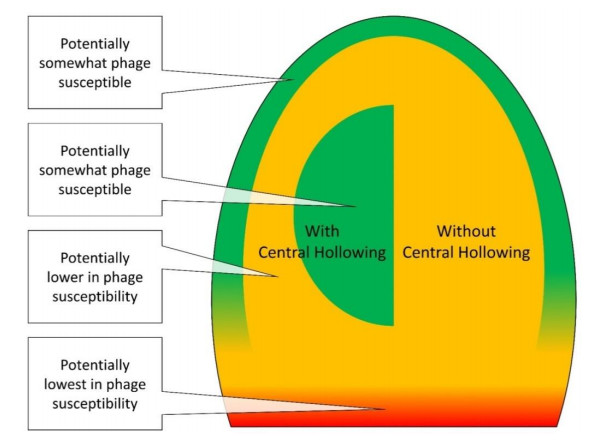

Figures(6)

Stephen T. Abedon. Phage “delay” towards enhancing bacterial escape from biofilms: a more comprehensive way of viewing resistance to bacteriophages[J]. AIMS Microbiology, 2017, 3(2): 186-226. doi: 10.3934/microbiol.2017.2.186







 DownLoad:
DownLoad: